Final Report for GNC07-079
Project Information
Conservation Reserve Program (CRP) restorations can benefit soil quality and fertility, but few evaluations of soil biological aspects of CRP lands exist. Measurement of soil microbial communities and activities may be useful in environmental evaluations of CRP prairie restorations. We examined microbial community composition under adjacent prairie- and corn-based agroecosystems, to test the general hypothesis that microbial community structure and activity would be distinct under varying vegetation and management treatments. Soil samples were taken in August of 2008 from two Conservation Reserve Program (CRP) prairie restorations and adjacent corn fields, on working farms in Dane County, WI. Principal component analysis (PCA) of the lipid biomarkers revealed soil microbial community structure within CRP prairie treatments displayed significant differences from the corn-based treatments. We found elevated abundances of arbuscular mycorrhizal fungi in the soils of prairie treatments, whereas under corn-based treatments we found elevated relative abundance of gram-positive bacteria, and increased ratios of gram-positive:gram-negative bacteria. We also found, in corn-based treatments, elevated production of cyclopropyl lipids compared to their precursor lipids, possibly indicating more physiological stress among bacterial communities in agricultural systems. These findings may reflect a more rhizosphere-dominated soil habitat within prairies, with greater microbial utilization of labile carbon sources, than in corn-based systems.
We suggest that the use of three types of lipid ratios, fungal:bacterial, Gm+:Gm-, and cyclopropyl/precursor, are most useful in distinguishing the effects of prairies from corn-based systems on soil microbial communities.
Introduction:
Preservation of soil quality, including soil fertility and structure, is a major goal of the growing number of public and private incentive-based ecosystem restoration projects. The U.S. Department of Agriculture’s Conservation Reserve Program (CRP) has provided incentives to landowners since 1985 to take environmentally sensitive land out of agricultural production and restore native vegetation. Much of this ‘sensitive’ land is classified as such due to high erodibility potential, such as steeply sloped land, or land especially prone to wind erosion. Some land is also enrolled in the program due to its suitability for wildlife habitat, protection of riparian areas, or other environmental benefits (Smith 2000). Currently, about 31 million acres are enrolled in the program, U.S.-wide (USDA 2009a). This comprises about 2% of land in the contiguous U.S., while cropland comprises about 19% of national land-use (USDA NRCS 2007). In the state of Wisconsin CRP-enrolled acres have decreased from a high of 700,000 in the mid-1990's to the 2009 level of approximately 461,000 (USDA 2009b).
The future of the CRP program is uncertain. Under the Food, Conservation, and Energy Act of 2008, total acres allowed to be maintained in CRP were reduced from 39.2 million acres to 32 million acres nationwide, beginning in fiscal year 2010. Some have advocated replacing the program with a harvestable biomass program in order to supply the U.S. with cellulosic bioenergy feedstocks for alternative energy projects. For example, in a U.S. Department of Energy assessment of potential U.S. acreage that could be dedicated to bioenergy production (U.S. DOE 2005), 50% of CRP biomass productivity (17-28 million dry tons) was assumed to be directed towards biofuel production, under the category of “idle cropland”. Many soil and climate scientists are advocating not for replacement of the CRP program, per se, but for expansion of governmental and market-based incentive programs to encompass prairie species-based restorations that could also potentially be used as harvestable cellulosic feedstock, and/or carbon sinks as part of a U.S. cap-and-trade program (Fargione et al. 2009).
Numerous studies have quantified the benefits of CRP restorations for wildlife habitat, soil quality and fertility, reduced soil erosion potential, and soil carbon accumulation (Gebhart et al., 1994; Staben et al. 1997, Baer et al. 2002; but, see Kucharik 2007). However, the biological activity of the soils in CRP restorations has largely been ignored. Staben et al. (1997) did quantify microbial parameters, including potential enzyme activities, in CRP soils, but did not assess microbial community composition. Soil microorganisms mediate important soil functions such as decomposition and cycling of nitrogen and phosphorus, and play a crucial role in development of soil structure. Additionally, recent evidence suggests that indicators of soil microbial community composition may be important predictors of soil processes and services, including carbon and nutrient cycling rates and dynamics (Smithwick et al. 2005, Carney et al. 2007).
Indicators of microbial community composition, in addition to total microbial biomass, may be suitable for assessing soil quality in CRP and other types of restoration sites. Biomarker compounds, such as phospholipid fatty acids (PLFAs), are one type of community structure indicator (Vestal and White 1989, Zelles 1999). Phospholipids are produced as part of microbial cell membranes, and are present in fairly constant amounts. Furthermore, their abundance reflects viable microbial biomass, as they are lost rapidly upon cell death (White 1993). Generally, PLFA data are useful in describing broad microbial community structure differences among different ecosystem and land-use types (Baath et al., 1995, Zelles et al. 1999, Burke et al. 2003).
In this study, we characterized soil microbial community composition and soil attributes within CRP prairie restorations and adjacent reduced-tillage cropland plots, on two working farms in south Dane County, Wisconsin. Our aims were: to determine whether CRP prairie restorations could affect soil microbial community composition; to determine whether soil carbon and soil aggregate-size distribution, important regulators of microbial communities, were affected by CRP restoration, compared to no-till corn management; and, to explore the use of microbial indicators that might differentiate between these two systems and be useful in future soil quality evaluations.
1) Evaluate the effects of CRP management on microbial biomass, soil microbial community composition and diversity. Relate microbial indices to soil quality parameters (especially soil carbon) that may respond to CRP and post-CRP cultivation practices.
2) Quantify soil carbon differences between paired CRP and cultivated sites. Examine the relationship between soil aggregates and soil carbon within CRP and cultivated sites.
3) Increase farmer knowledge of soil quality, and increase the ability of farmers to preserve long-term soil quality through dissemination of educational materials and decision-making tools.
Cooperators
Research
Samples were collected from two private farms in southwestern Dane County (here designated Site A and Site B). The sites were within 5 km of each other to ensure similar climatic conditions. Both sites contained “treatment” plots of mature CRP prairie restoration paired in space with treatment plots of corn cultivation. At Site A, the prairie restoration was 5 years old and located on a north-facing slope. Management of the prairie included a controlled burn in spring 2008. The plot of corn cultivation at Site A was in a 2-year rotation with soybeans for 10 years, and had not been tilled during this time. At Site B, the CRP prairie restoration was over 10 years old, while the plot of corn cultivation was in its first year of cultivation after having previously been part of the CRP restoration. The corn had been planted under no-tillage conditions and was preceded in the season by tilled winter wheat.
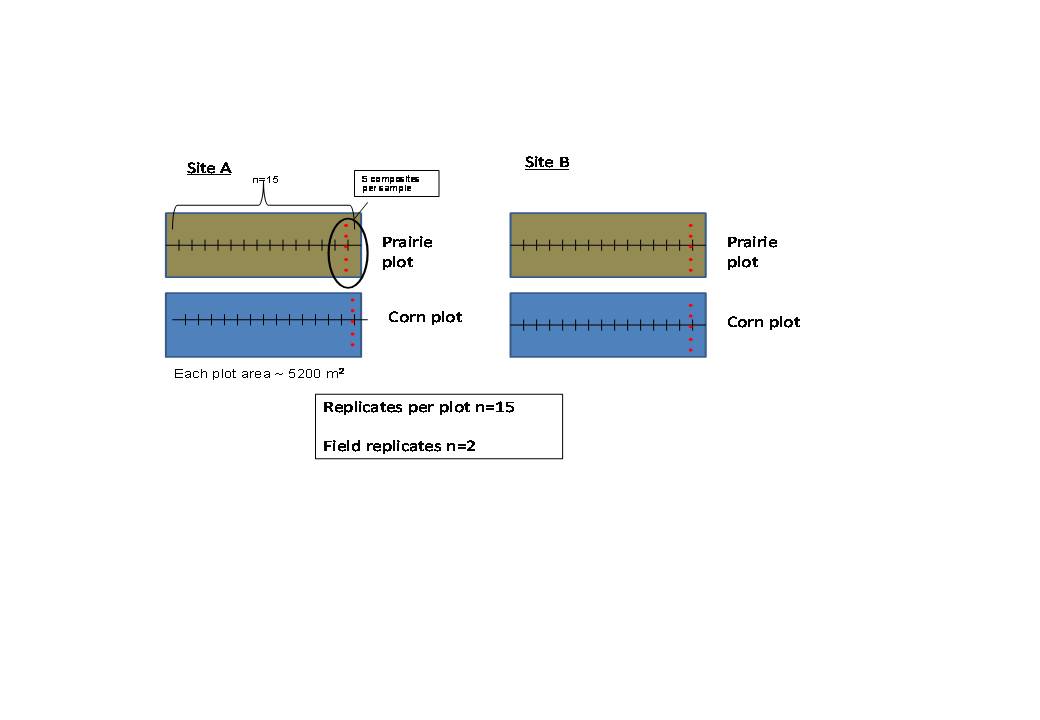
Sampling was conducted during one week in August 2008, following plant peak growth. At each plot (one prairie and one cornfield at each site), we sampled an area of approximately 5200 square meters (Figure 1). Fifteen soil samples per plot were taken along a 450 meter transect, each sample made up of five composite cores in order to reduce sampling variability. Each core for compositing was taken with a push probe of 5 cm diameter to a depth of 25 cm. Cores were broken into two depths, 0-10 cm, and 10-25 cm. We also took ten bulk density cores per plot, along the same transect.
Following sampling, soils were transported immediately to the laboratory, where we removed large roots (> 4 mm) and rocks, and split the samples into two subsamples. We homogenized and froze one set of subsamples at -20º C for microbial analysis, and another subset was kept in plastic bags in a field-moist state until physical fractionation one week later.
We analyzed each sample for total carbon and total nitrogen analysis using a Elementar VarioMax CN analyzer (Elementar, Hanan, Germany). We measured soil pH by 1:1 determination of air-dried soil in deionized water (Thomas 1996). We measured soil texture by particle size analysis by the hydrometer method. We obtained soil water content gravimetrically by drying soil at 110º for 24 hours and quantifying water loss.
We measured soil microbial community composition from bulk soils in each treatment using membrane lipid analyses, as described by Smithwick et al. (2005). Since phospholipids in cell membranes are metabolized rapidly upon cell death, phospholipid analysis of soil microorganisms provides an estimate of viable microbial biomass White et al., 1979).
Briefly, we extracted, purified, and identified cell membrane lipids using a PLFA extraction based upon a modified Bligh and Dyer (1959) technique, followed by modified FAME analysis (Microbial ID Inc., Hayward, CA, USA). Lipids were extracted from 2.5 g of freeze-dried, homogenized soil using a chloroform-methanol extraction with a phosphate buffer of pH 7.0, in a volume ratio of 1:1:2 buffer:chloroform:methanol. Extracted samples were subjected to saponification, methanolysis, and methylation to cleave fatty acids from cell lipids and convert the resulting fatty acids to fatty acid methyl esters, and then a final extraction to an organic phase. Subsequently, we analyzed 2 ?l aliquots of the fatty acid methyl esters (FAMEs) using a Hewlett-Packard 6890 Gas Chromatograph equipped with a flame ionization detector and an Ultra 2 capillary column (Agilent). We identified lipid peaks using bacterial fatty acid standards and Sherlock peak identification software (Sherlock microbial identification system, MIDI Inc., Newark, DE, USA). Response peak areas were converted to ?mol lipid g-1 soil using internal standards 9:0 (nonanoic methyl ester) and 19:0 (nonadecanoic methyl ester). Internal standard peak areas were subtracted from final response peak areas prior to statistical analysis. Lipids present at less than 0.5 mol % (unless present in a majority of samples from each treatment) were excluded from further analysis.
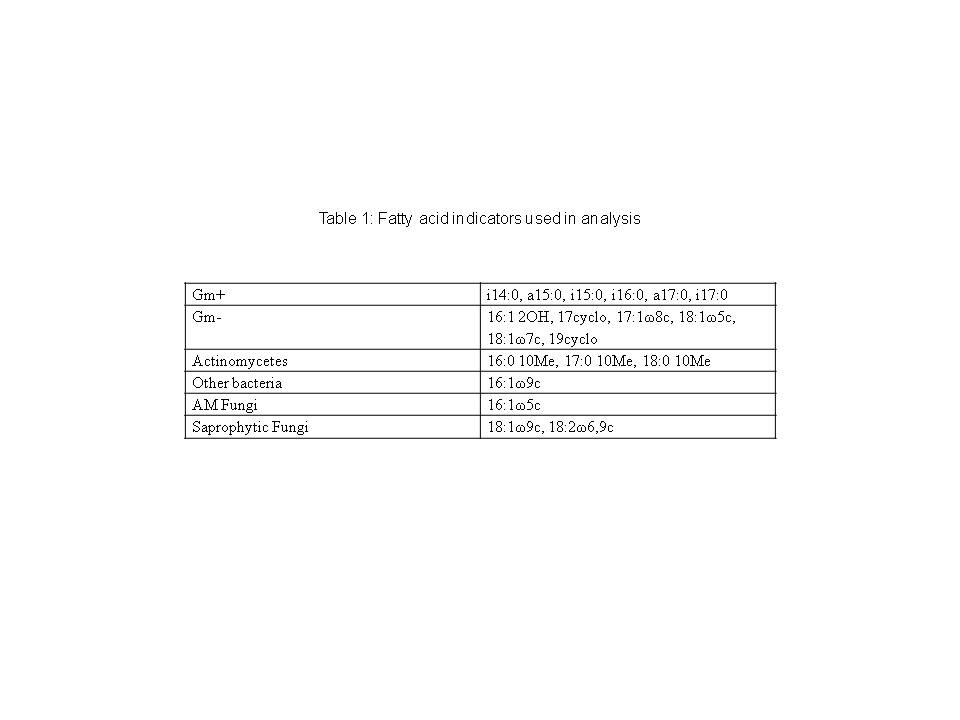
We used the complete set of lipids generate microbial ‘fingerprints’ by Principal Components Analysis, and focused on a smaller set of indicator lipids for ecological guilds of interest. These are summarized in Table 1. Generally, gram-positive bacteria can be represented by the branched lipids: i14:0, a15:0, i15:0, i15:1, i16:0, a17:0, i17:0. Gram-negative bacteria are indicated by mono-unsaturated and hydroxy lipids: 16:1 2OH, 17:0cy, 17:1?8c, 18:1 ?5c, 18:1 ?7c, and 19:0cyclo (Ratledge and Wilkinson 1988, Bardgett et al. 1996, Wilkinson et al. 2002, Santruckova et al. 2003, Steenwerth et al. 2003, Kaur et al. 2005, Mentzer et al. 2006b, Zelles 1997). Arbuscular mycorrhizal fungi (AMF) are indicated by 16:1?5c and saprophytic fungi (SF) by the sum of 18:1?9c and 18:2?6c (Balser et al. 2005).
Fatty acid nomenclature is described as “A:B?C” where “A” is the total number of carbon atoms, “B” is the number of double bonds, or unsaturations, and “C” is the position of the double bond relative to the methyl end of the molecule. Prefixes “i” and “a” refer to iso and ante-iso methyl branching. Hydroxy groups are indicated by “OH” while cyclopropyl groups are denoted by “cyclo” (Steenwerth et al., 2003). We measured total PLFA as an indicator of total microbial biomass, total biomass of microbial groups and individual lipids (nmol lipid g-1 dry soil), as well as relative abundance of the above groups (mol %). The measure of relative abundance expresses the proportion a lipid makes up of the total microbial biomass of a sample.
To better assess the relationships between microbial community composition and soil structure among different treatments, we chose a subset of samples from each treatment to fractionate into different sized soil aggregates. Aggregates were separated by size according to a modified wet-sieving procedure (Elliott 1986), also described in Simpson et al (2004). We performed a physical fractionation by wet-sieving of 24 fresh samples (six per plot), into macroaggregates (> 250 µm), microaggregates (53 µm-250 µm), and the silt and clay fraction (<53 µm). Due to concerns that sieving with water would lyse microbial cells and thus preclude microbial analysis, we used a solution of 14.5 mM CaSO4. One week after sampling, a 25-gram subsample of homogenized soil was immersed in a 14.5 mM CaSO4 solution on a 250 µm sieve for five minutes. We then moved the sieve up and down 5 cm vertically by hand for 50 times within two minutes. We collected those aggregates retained by the 2 mm sieve by hand and then washed those not retained onto a 53 µm sieve. The procedure was repeated, and each fraction was collected and frozen at -20º C immediately. Frozen fractions were then freeze-dried and homogenized. We analyzed each sample for total carbon and total nitrogen analysis using an Elementar VarioMax CN analyzer (Elementar, Hanan, Germany). From four of the freeze-dried samples (one per plot), we extracted microbial lipids in the same manner as detailed above. We extracted lipids from 2.5 grams of the macroaggregate fractions. We used 0.5 grams of the microaggregate fractions due to the small mass of microaggregate sample available following fractionation.
To test whether fractionation of frozen or fresh soils affected subsequent lipid analysis, we additionally performed the same physical fractionation and lipid extraction procedure on soils that had been frozen at -20º C and thawed over 48 hours at 4º C (to avoid possible disruption of the microbial community due to rapid temperature changes). It was determined that microbial community structure did not differ significantly between frozen or fresh samples (data not shown); the fresh sample values were used for all subsequent analysis.
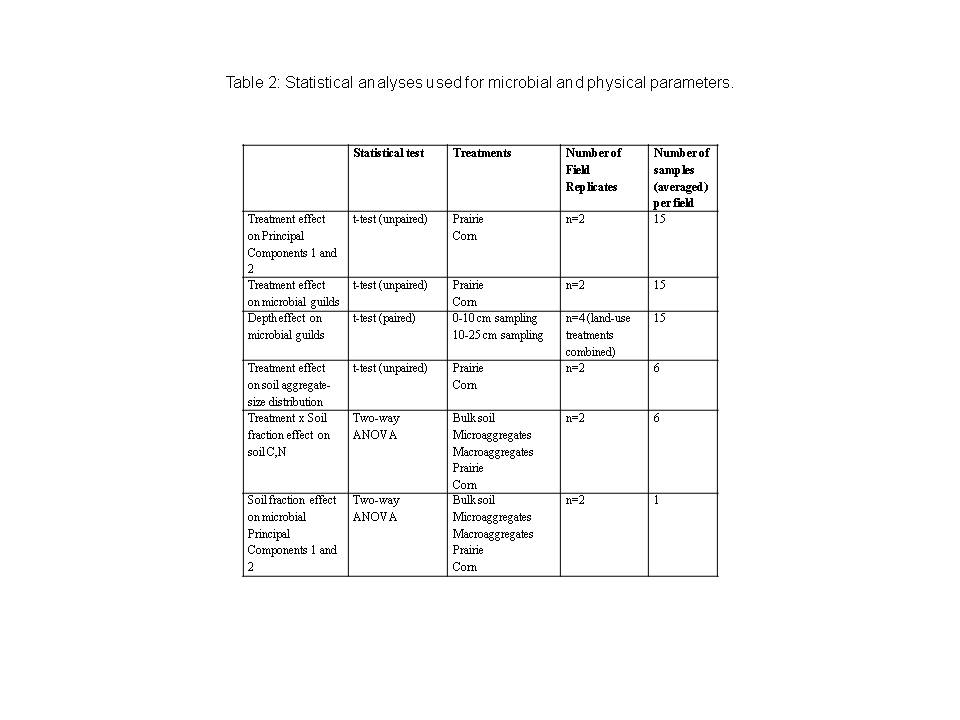
We used principal component analysis on the arcsin transformed mol fraction (relative abundance) of lipids to provide a general microbial lipid ‘fingerprint’ (JMP, SAS Institute, 2002) of each treatment and depth sampled. An independent sample t-test on the principal components (PC1 and PC2) was conducted to assess the effects of treatment on overall microbial community structure. To assess possible treatment effects on physico-chemical response variables, and those of total microbial biomass, individual lipid biomass and relative abundance, microbial guild biomass and relative abundance), we used unpaired t-tests of the means of prairie and corn sites (n=2). To assess differences in depth on these response variables, we used paired t-tests between the 0-10 cm and 10-25 cm depths of all treatments (n=4). Table 2 summarizes the statistical analyses we utilized.
We also tested the effects of soil fraction on microbial and physical variables (Table 2). We used two-way analysis of variance (ANOVA) to test the effects of treatment by soil fraction for total carbon and total nitrogen, and an unpaired t-test of the effects of treatment on % macroaggregates, % microaggregates, and % silt and clay fraction. For tests of the effect of soil fraction on microbial community structure, we also used principal component analysis on the arcsin transformed mol fraction (relative abundance) of lipids. We conducted two-way ANOVA on the principal components PC1 and PC2 to determine the effects of soil fraction and land-use treatment on overall microbial community structure. All ANOVA tests were followed by Tukey’s post-hoc analysis for pairwise comparisons (JMP, SAS Institute 2002).
{kind=link}
{kind=link}
{kind=link}
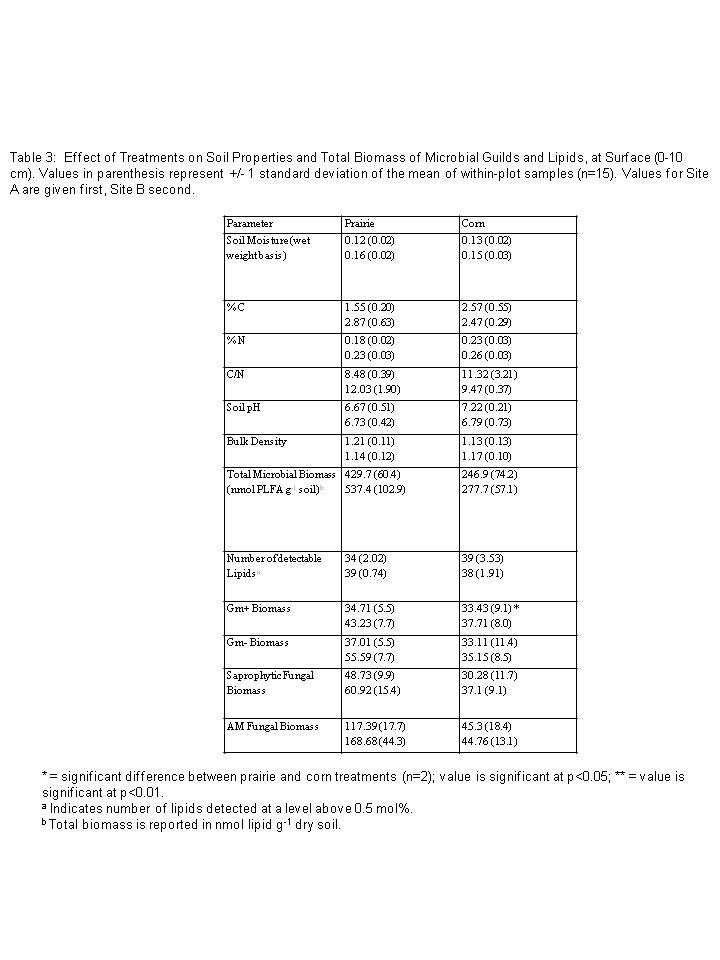
The physical and chemical soil properties of corn and prairie soils were very similar (n=2, Table 3). Total carbon, nitrogen, or C:N ratio were not significantly different between bulk soils in corn and prairie treatments. Bulk density and pH values between the two treatments were also similar (Table 3). Table results report values for Sites A and B separately due to the variation in soil and microbial properties across the two sites.
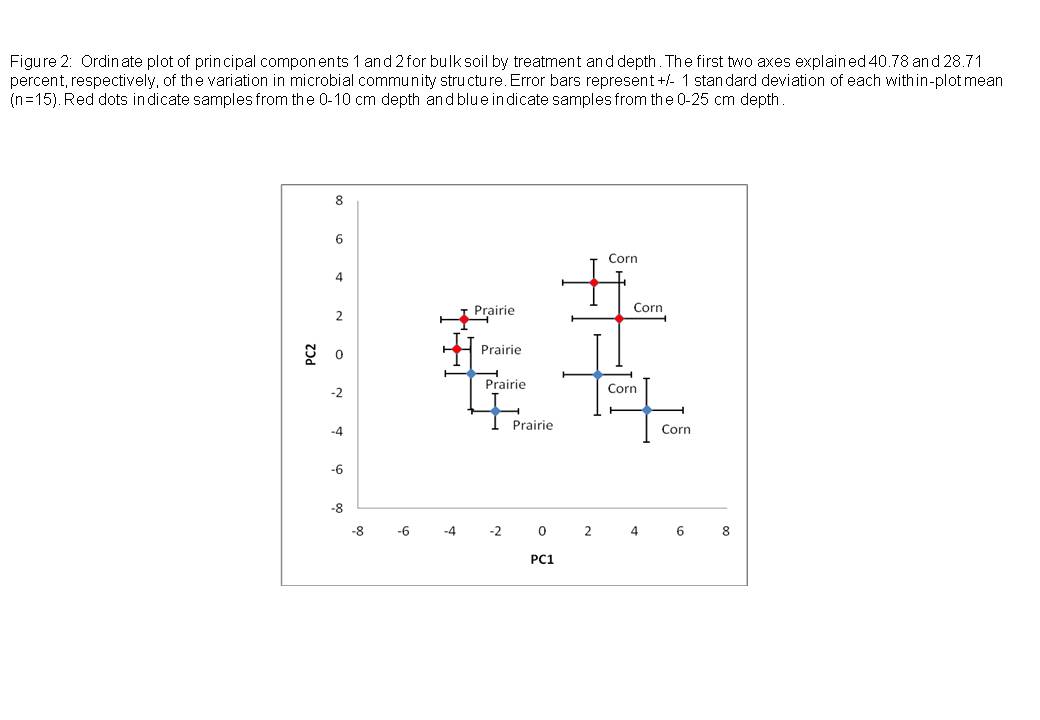
Total microbial biomass tended to be higher in prairie plots than in corn plots (Table 3). Principal components analysis (PCA) of arcsine transformed mol percent data also showed significant differences in overall microbial community structure between prairie and corn treatments (Figure 2). A t-test on average PC1 values showed that microbial community composition was significantly different between prairie treatments and corn treatments (p<0.01) along the first axes (PC1). PC1 and PC2 explained 40.78 and 28.71 percent, respectively, of the variation in microbial community structure.
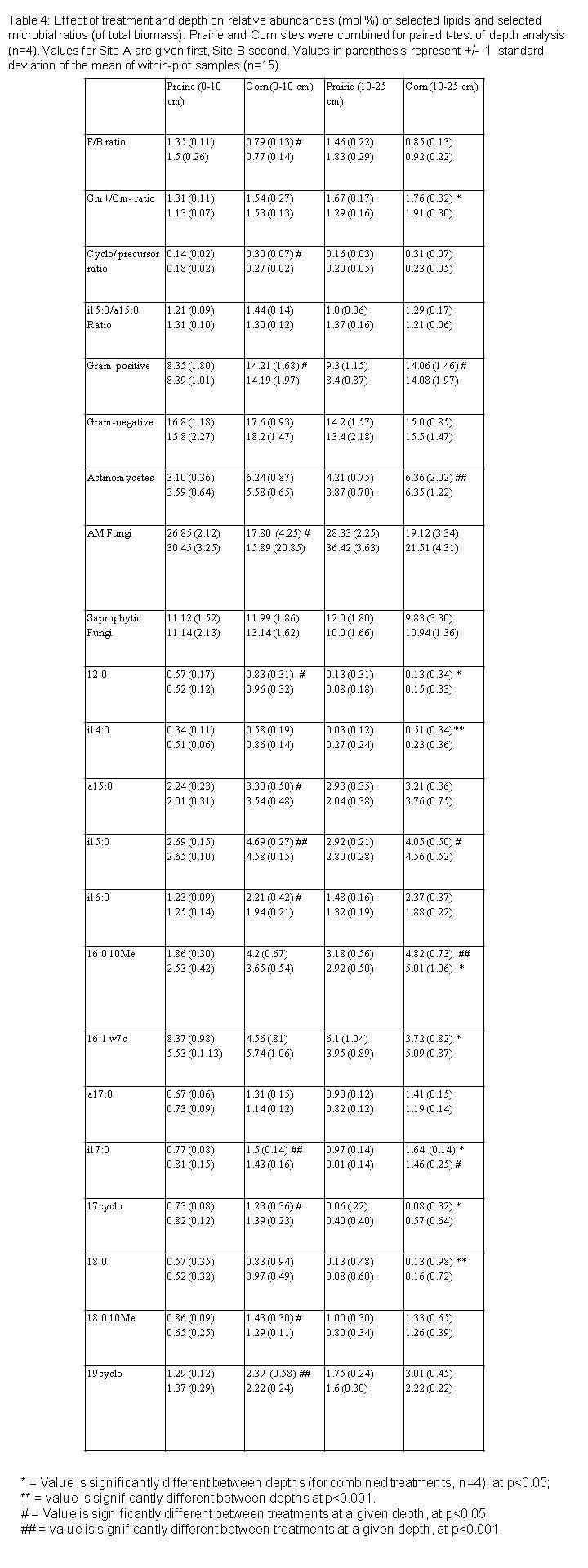
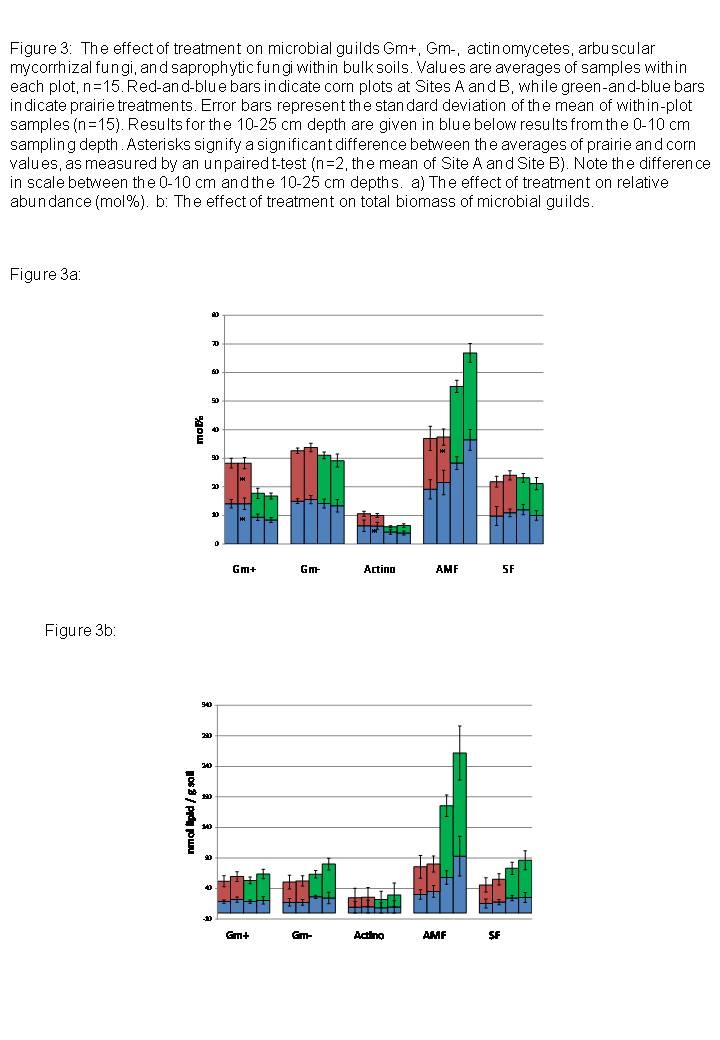
In the 0-10 cm layer, the relative abundance (mol %) of Gm+ bacteria in general was significantly greater in the corn plots than in prairie (Figure 3a, p<0.05). The relative abundance of several specific Gm+ lipid indicators (a15:0, i15:0, i16:0, a17:0, i17:0, as well as the Gm- indicator 19cyclo) was likewise greater in corn plots than prairie (Table 4). The relative abundance of AM fungi, by contrast, was significantly greater in prairie plots than corn (p<0.05, Table 4, Figure 3a), while the relative abundance of saprophytic fungal markers did not show significant differences between treatments (Table 4, Figure 3). Although actinomycetes are members of the Gm+ bacterial group, we measured them as a distinct group. The relative abundance of actinomycete markers tended to be higher in corn treatments but this difference was not statistically significant at the 0-10 cm layer, only at the 10-25 cm layer (p<0.05, Figure 3).
On average, total biomass of Gm+ bacteria, Gm- bacteria, actinomycetes, saprophytic fungi, or AM fungi, did not differ significantly between corn and prairie treatments (Figure 3), in either the 0-10 cm layer or the 10-25 cm layer. Saprophytic fungal biomass tended to be higher in corn plots, while AM fungal biomass tended to be higher in prairie plots. However, total fungal biomass varied greatly by site, and the mean biomass values were not statistically significant at a sample size of two (Site A and Site B).
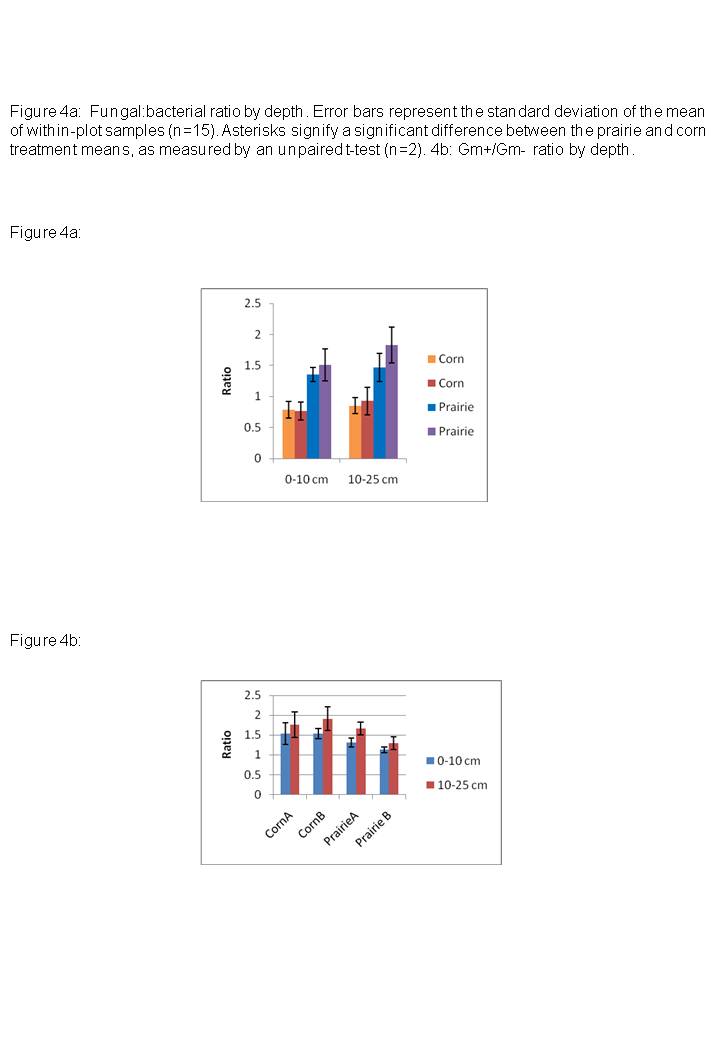
We measured several ratios to further examine treatment differences on microbial parameters. The ratio of cyclopropyl lipids (17cyclo + 19cyclo) to their monoenoic precursors (16:1?7c + 18:1?7c), often used as an indicator of Gm- nutritional stress (Guckert et al. 1986, Kieft et al. 1994, 1997), was significantly higher in the corn plots than in the prairie (Table 4); however the ratio of i15:0/a15:0, a common indicator of Gm+ stress, was not significantly different between treatments. The ratio of fungal/bacterial lipids was also significantly higher in the prairie plots than the corn plots, in the 0-10 cm layer (p<0.05) but not in the 10-25 cm layer (p<0.10, Table 4, Figure 4a).
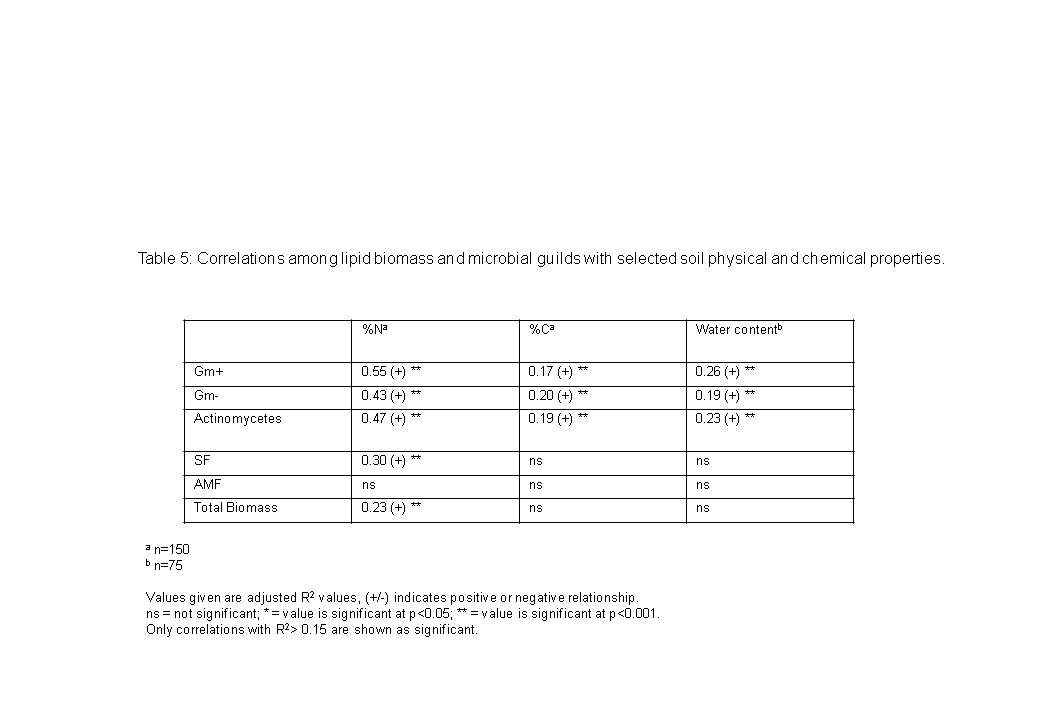
To relate soil microbial community composition with soil habitat parameters, we correlated several soil physico-chemical properties (total C, total N, and water content) with lipid biomass and total biomass of microbial guilds (Table 5). Total N tended to correlate most highly (and positively) with microbial groups, while total C did not correlate as well. Bacterial guilds tended to be more highly correlated to all of these properties than fungal markers were (Table 5).
The total biomass of most guilds and lipids declined with depth, as did total microbial biomass (data not shown). Microbial communities from the two depths studied (0-10 cm and 10-25 cm) displayed a moderate degree of separation along the PC2 axis (Figure 2), as indicated by Principal components analysis. A paired t-test of PC2 values from the two depths indicated that this separation was statistically significant (p<0.01). The relative abundance (mol %) of most guilds and lipids did not vary with depth, but for several, including i14:0 (Gm+), 16:1w7c (Gm-), and 17cyclo (Gm-), the relative abundance declined significantly with depth (Table 4), while several lipids, including the generically produced Gm+ lipid, 19:0, Gm+ lipid i17:0, and the Gm- indicator 19cyclo, tended to show moderate increases in relative abundance (Table 4).
Paired t-tests that included all sites (n=4) showed that the ratio of Gm+/Gm- lipids was slightly higher at the 10-25 cm depth than at the 0-10 cm depth (p<0.05; Table 4, Figure 4b). Neither the ratio of cyclopropyl lipids to their precursors, nor the F:B ratio differed significantly between depths within either treatment (Table 4, Figure 4a).
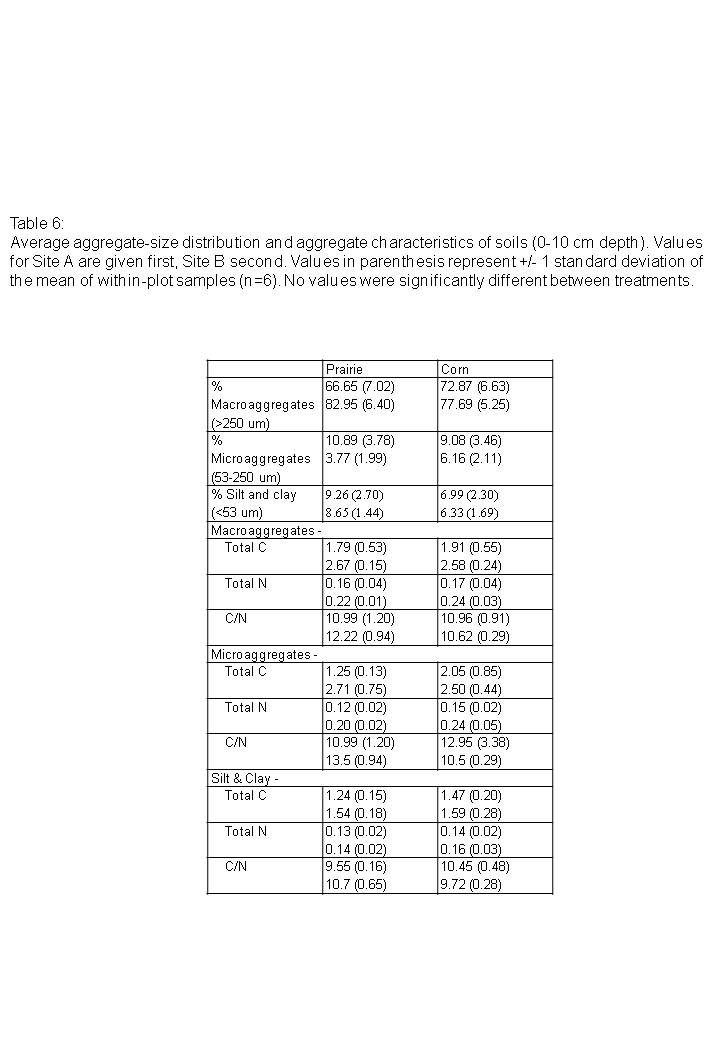
No-till corn and prairie treatments were very similar in aggregation characteristics (Table 6). The proportion of macroaggregates, microaggregates, and the silt and clay fraction did not vary with corn or prairie treatment. Land-use did not affect the proportion of carbon or nitrogen located within macroaggregates, microaggregates, the silt and clay fraction, or within bulk soil.
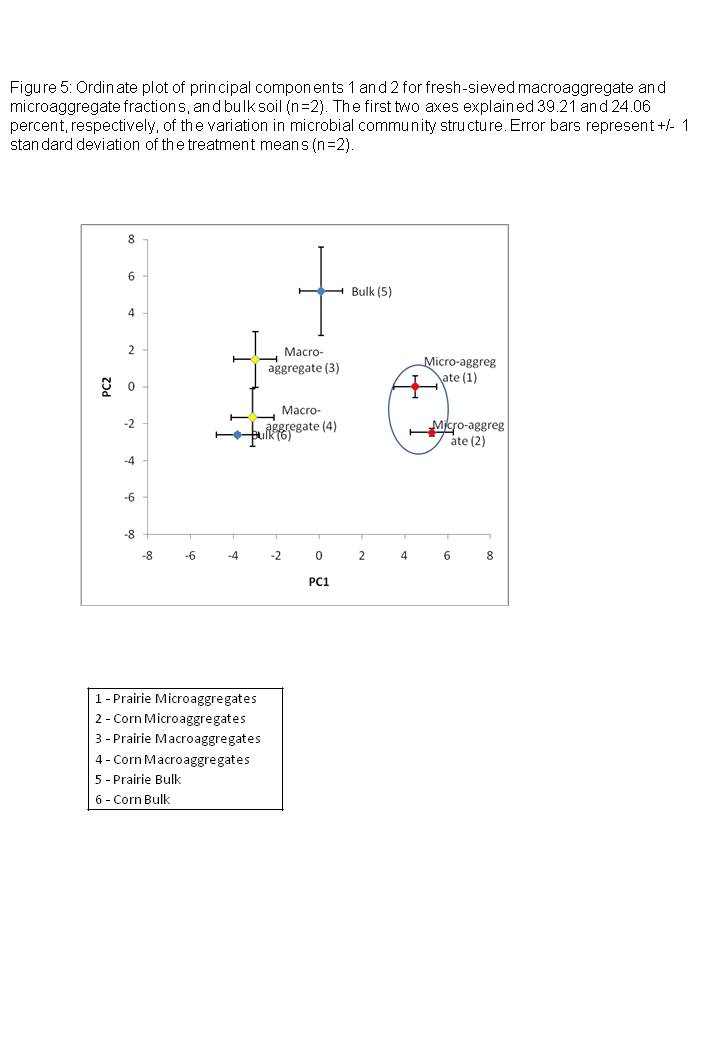
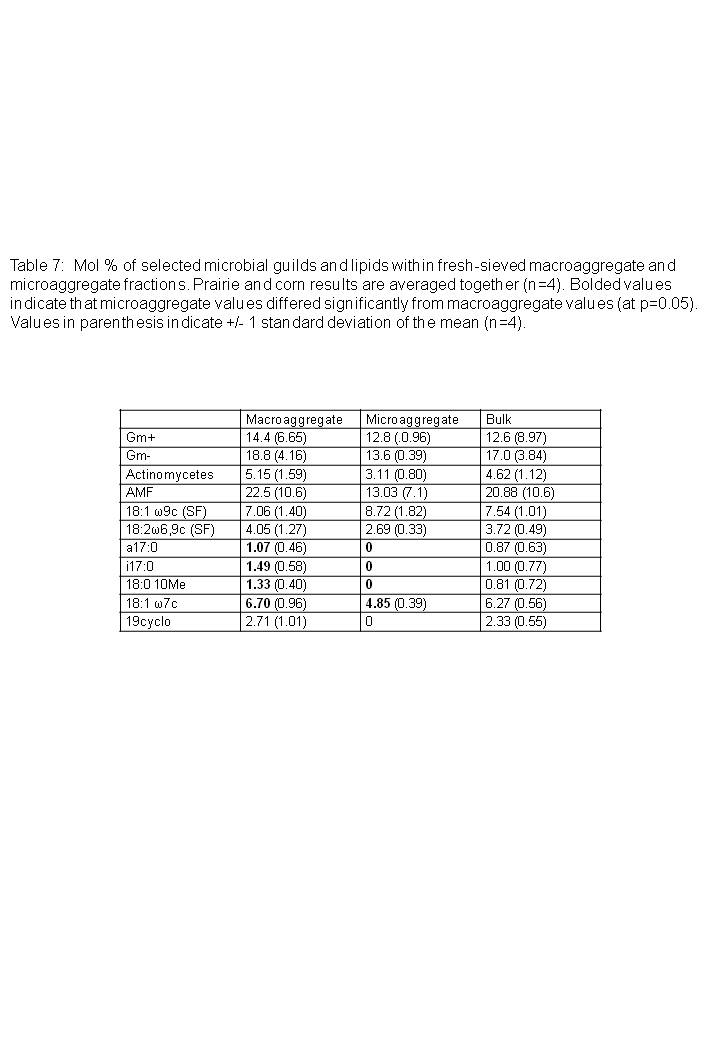
Principal components analysis showed that microbial community structure differed between microaggregate fractions, macroaggregates, and bulk soils (Figure 5). The first two axes explained 39.21 and 24.06 percent, respectively, of the variation in microbial community structure. ANOVA conducted on PC1 showed that community structure in microaggregates differed significantly from macroaggregates and bulk soils (p<0.01, Figure 5), while bulk soils and macroaggregate fractions were similar in structure. Fractions were not significantly different along the PC2 axis, although several of the branched and monounsaturated lipids were not present at all in the microaggregate fractions but were present in macroaggregate and bulk fractions (Table 6). The proportion of saprophytic fungal markers did not vary among any of the size classes. Finally, the proportion of AM fungal marker 16:1?5c was lower in the microaggregate portions, although the differences were not statistically significant.
Our results indicate that CRP prairie restorations can substantially affect soil microbial community properties. Overall microbial community structure differed substantially between prairie restorations and adjacent corn plots. This was the case for both Site A and Site B, despite significant differences in land-use history between the two sites; Site B had only been removed from the CRP restoration and planted to corn for one year, whereas both the treatments at Site A had been planted for a number of years. Additionally, the relative abundances of important microbial functional guilds, such as arbuscular mycorrhizal fungi, shifted dramatically between prairies and corn plots, suggesting that prairie restorations can be effective in changing soil biological properties even though basic physico-chemical properties of the sites remained unaffected. We suggest that the total microbial biomass, the fungal:bacterial ratio, and the proportion of AM fungi, are particularly important indicators to measure in determining soil quality in ex-arable or reclaimed land. This is in line with calls from other restoration ecologists (Harris 2009.
We found a trend toward higher overall microbial biomass within CRP plots compared to the no-till corn plots. This concurs with previous research; for example, Staben et al. (1997) found that CRP prairie restorations increase the total biomass and microbial activity of the soil. Since microbial biomass generally correlates well with carbon inputs into the soil system, it is likely that greater microbial biomass in CRP restorations reflects the effects of dense, perennial vegetation providing greater aboveground litter. Prairie systems are also more likely than corn systems to have dense, fine root systems, thus likely exuding greater amounts of carbon substrate to the soil through the root, and elevating the total microbial biomass by boosting populations in the rhizosphere.
It is interesting to reflect on how soil microbial biomass might respond to potential alternative management scenarios of perennial crops. For example, if perennial systems are increased on the landscape in order to provide biofuel feedstock, they will likely be harvested one or two times per year, and nitrogen fertilization will likely be used extensively to boost yield. Harvest of perennial crops would substantially decrease aboveground litter inputs of substrate to microbial populations, but root inputs would remain intact. N fertilization, on the other hand, may enhance aboveground growth at the expense of belowground growth, impacting the root:shoot ratio of growing perennial plants, and potentially reducing belowground inputs of carbon to the microbial community (Heggenstaller et al. 2009). More research is needed on the belowground impacts of these novel management activities in prairies.
We had expected to find indicators of bacterial nutritional stress within corn plots, compared to prairie restorations. Our results indicated that corn treatments tended to have higher cyclopropyl/precursor ratios than prairies, suggesting higher nutritional stress in Gm- bacterial populations in the corn treatments. Previous researchers have postulated that high cyclopropyl/precursor ratios indicate a bacterial community response to carbon limitation, possibly a high C:N ratio (Fierer et al. 2003, Allison et al. 2005). Although our treatments did not significantly differ in soil C:N, it is likely that plant provision of labile carbon into the rhizosphere differs between treatments. We suggest that instead of total C being a limiting factor to microbial communities in this environment, labile C stocks may differ due to differences in vegetative inputs and differences in C quality in these systems. Root exudations and fine root turnover impart considerable quantities of labile, low-molecular weight carbon compounds to the soil. We suggest that a rhizosphere-oriented soil habitat within the prairie restorations decreases indicators of Gm- nutritional stress, whereas Gm- bacteria in cultivated plots experience physiological stress due to inadequate quantity, or quality of labile carbon compounds. Indeed, Staben et al. (1997) found that in CRP land, compared to wheat-fallow systems, total carbon did not differ between land-uses, but carbon pools and C mineralization potential was significantly greater in CRP treatments, suggesting microbial discrimination between types of carbon. This may be the case even when aboveground shoot residues are plentiful, as in no-till corn systems, due to potentially low litter quality as a microbial substrate.
We found a strong effect of prairie restoration on AM fungal relative abundance in both sites. This was the case even though at Site A, the corn treatment had not been tilled in over 5 years. This suggests that, although tillage can be a dominant control on fungal communities, other factors may also be at work here in suppressing fungal abundance in cultivated sites. For example, nitrogen additions have also been shown to reduce fungal abundance (de Vries et al. 2007). Others have also found that relatively young restored prairies can have high fungal:bacterial ratios (Allison et al. 2005) relative to cultivated systems. According to Allison et al. (2005), the F:B ratio increases rapidly within the first few years of restoration, leveling off thereafter. They found that in older prairies (for example, those >25 years old) the F:B ratio actually decreased due to increased bacterial biomass. It is possible that as prairie restorations age, they develop different biological properties than earlier in the restorations. Nevertheless, our results confirm that in prairies of 5-10 years of age, the dominant microbial indicator of restoration appears to be the resurgence of arbuscular mycorrhizal fungi, especially relative to bacterial biomass.
Our results did not show evidence that aggregate size distribution differed between the no-till cultivated systems and the prairies. This may indicate that when maintained, no-till management can be as effective as prairie restorations in restoring certain aspects of soil physical quality, such as large macroaggregates. However, because we did not measure baseline aggregate-size distributions before the land-use treatments were implemented, it is possible that the sites’ aggregate size distributions may reflect underlying site differences rather than the influence of land-use. Additionally, we only measured proportion of macroaggregates, free microaggregates, and the silt and clay fraction; it is also possible that there are finer differences in aggregate sizes (such as variations in size of macroaggregates) that may vary with land-use.
Due to resource and time limitations, we had a small number of samples for microbial community composition in differently sized soil aggregates. Nevertheless, our study serves as a pilot study on the use of PLFA-FAME analysis on the ecologically relevant unit of the soil aggregate. Results suggest that microbial community composition may differ between different types of soil spatial units, specifically micro- and macroaggregates, and that soil aggregation characteristics may be important in structuring microbial communities. Our results indicated that microaggregate microbial communities displayed greater similarity to each other than to macroaggregate or bulk soils from the same land-use. We suggest that because microaggregates are more stable than macroaggregates (Oades 1984), and form an isolated environment for microorganisms within, microbial communities and activities within microaggregates may be more insulated from the effects of land-use than are macroaggregates. Variation in microbial communities with aggregate size suggests that environmental factors, including land-uses such as tillage-based systems, that affect aggregate size distribution may contribute to shifts in the composition of microbial communities.
Previous studies of microbial communities in soil size fractions have also found differences among fractions (Schutter and Dick 2002, Blackwood and Paul 2003). There are several plausible reasons why microbial communities may differ between macroaggregates and microaggregates. Differences between micro- and macroaggregates may be caused by differences in resource provision between the two different size classes. Research has shown that macroaggregates contain organic matter that is more labile, or less decomposed, than that in microaggregates (Elliott 1986, Oades and Waters 1991). This could provide a mechanism whereby microbial community members with different carbon substrate preferences, or abilities, persist in differently-sized fractions. Additionally, microorganisms contained within microaggregates may be better protected from attack by protozoa or other soil predators (Elliott et al. 1980, Six et al. 2006), so particularly vulnerable microorganisms may be disproportionately located within microaggregates. More research is needed to elucidate which mechanisms are most important in determining the spatial location of different types of microorganisms.
Our study did not include sufficient replication (our sample size was small, n=2) for drawing substantial conclusions about differences in the biological properties of aggregate sizes. A further limitation was that we examined only ‘free’ microaggregates; we did not further separate the fractions from macroaggregates and microaggregates. In other words, we have not been able to distinguish between free microaggregates, which we looked at, and microaggregates that may be occluded within macroaggregates. Further separation may yield different results for microbial community structure or distribution of carbon (Denef et al. 2004). However, our results seem to indicate that future research into the interactions between different members of the microbial community and aggregates of various sizes is warranted.
Our sites (A and B) were chosen for their proximity to one another, for their status as working farms, thus representative of, and relevant to, real conditions on the landscape, and also because they both contained CRP/CREP prairie restorations adjacent to corn cultivation. However, sites A and B exhibited considerable differences in environment and management from each other, and presumably from other farms in the region. Therefore, our inferences only extend to the particular study site, and generalizations about the larger landscape are limited.
Although the results of principal components analysis demonstrated differences between restorations and corn treatments, further study into the mechanisms structuring microbial communities is required, in order to elucidate which environmental factors contribute to differences in microbial community structure. Snapshot studies such as the current one can be useful as exploratory analyses or for generation of baseline data. But long-term studies that record variability and trends in microbial communities, and other potential soil quality indicators, over time, are needed to assess the success of restorations in affecting soil microbial communities.
- Effect of treatments on soil and microbial properties
- Effect of treatment and depth on microbial abundance
- Effect of aggregate size on microbial community structure
- Soil aggregation characteristics
- Effect of aggregate size on microbial guilds
- Effect of treatment and depth on microbial community structure
- Effect of treatment on microbial guilds
- Fungal:bacterial ratio and Gm+:Gm- bacterial ratio
- Correlations among microbial biomass and soil properties
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Educational & Outreach Activities
Participation summary:
- Lipps, Laura A. 2010. Evaluation of corn-based and perennial bioenergy cropping systems: Effects on soil microbial communities. M.S. Thesis. University of Wisconsin-Madison. Lipps, L.A. and Balser, T.C. 2010. Effects of bioenergy agroecosystems on soil microbial communities. Wisconsin Integrated Cropping Systems Trial 12th Annual Report. Public Presentation: Lipps, L.A. 2010. Evaluating bioenergy crops: crop effects on soil microbial communities. University of Wisconsin-Madison Nelson Institute seminar.
Project Outcomes
Many farmers in Wisconsin may be facing financial incentives to remove land from the CRP. Our results suggest that even if basic soil test parameters (such as pH, total carbon, and nitrogen) are not demonstrably changed by conversion of CRP land to corn cropland, soil microbial community composition may be affected, especially the ratio of AM fungi to bacterial biomass. Shifts in the abundance or dominance of AM fungi may have important consequences for soil quality. They have been shown to contribute substantially to the formation of macroaggregates in soils by binding soil particles together with fungal hyphae, and by production of the binding agent glomalin (Rillig et al. 2002, Hallett et al. 2009). AM fungi can also contribute substantially to plant productivity, especially in infertile soils, by increasing plant acquisition of soil nutrients (Wilson and Hartnett 1997).
Our results suggest that no-till management of corn may not significantly alter physico-chemical measures of soil quality enough to be detrimental to soil functions. This is in contrast to conventionally tilled soils, which have been found to have lower soil organic carbon as well as lower measures of soil aggregation (Denef et al. 2004). The lack of difference in soil physico-chemical parameters between corn and prairie treatments does not necessarily suggest equivalence of treatments on soil quality, because of underlying soil and landscape factors that might also be influencing the soil habitat. Many studies register gains in soil quality through participation in the CRP, and our results are not incompatible with those findings. However, several recent studies have cast doubt on the conventional wisdom that soil quality, and soil carbon sequestration, is necessarily improved by CRP prairie restoration (Brye et al. 2002, Kucharik 2007). Effects of CRP management, compared to cropland on the similar soil type, may be strongly affected by plant species composition and management practices such as burning or control of invasive and woody species (Kucharik 2007). On a similar note, restored prairies, even decades after restoration, have been found not to have recovered essential ecosystem functions and services that native prairies provide, and soil microbial community profiles remain distinct (McKinley et al. 2005). These cautionary studies suggest that microbial community composition, as well, may display variable responses to CRP restorations. This may be especially the case if future CRP prairies are managed in diverse ways, for instance, if prairie plantings are harvested for cellulosic biomass, or fertilized to boost aboveground yield. Future managers of perennial systems, whether they include prairie restorations for aesthetic purposes, governmental incentives, biomass production for harvest, or carbon sequestration, should be aware that yearly follow-up management of restorations to specifically favor soil quality may be necessary to achieve positive environmental benefits.
In conclusion, we found significant differences between soil microbial communities within CRP prairie restorations and adjacent reduced-tillage cornfields. In particular, microbial communities could be differentiated by the fungal:bacterial ratio, and the proportion of Gm+ and AM fungi, as well as elevated cyclopropyl/precursor ratios in corn plots. These may be particularly important indicators to measure in determining effects of land-use on soil quality. As a next step, the functional role of these microbial groups in processes important to soil quality, such as carbon dynamics, nutrient mineralization, and soil aggregation, should be further investigated.
Allison, V.J., R.M. Miller, J.D. Jastrow, R. Matamala, and D.R. Zak, D.R. 2005. Changes in soil microbial community structure in a tallgrass prairie chronosequence. Soil Science Society of America Journal 69, 1412–1421.
Bååth, E., A. Frostegård, T. Pennanen, and H. Fritze. 1995. Microbial community structure and pH response in relation to soil organic matter quality in wood ash fertilized, clear-cut or burned forest soils. Soil Biology and Biochemistry 27, 229–240.
Baer S.G., D.J. Kitchen, J.M. Blair, and C.W. Rice. 2002. Changes in ecosystem structure and function along a chronosequence of restored grasslands. Ecological Applications 12, 1688–1701.
Balser, T.C. and M. Firestone. 2005. Linking microbial community composition and soil processes in a California annual grassland and mixed-conifer forest. Biogeochemistry 73, 395–415.
Balser, T.C., K.K. Treseder, and M. Ekenler. 2005. Using lipid analysis and hyphal length to quantify AM and saprotrophic fungal abundance along a soil chronosequence. Soil Biology and Biochemistry 37, 601-604.
Bardgett, R. D., P.J. Hobbs, and Å. Frostegård. 1996. Changes in fungal:bacterial biomass ratios following reductions in the intensity of management on an upland grassland. Biology and Fertility of Soils 22, pp. 261–264.
Blackwood, C.B., and E.A. Paul. 2003. Eubacterial community structure and population size within the soil light fraction, rhizosphere, and heavy fraction of several agricultural systems. Soil Biology and Biochemistry 35, 1245–1255.
Bligh, E. G., and W. G. Dyer. 1959. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology 37, 911–917.
Burke, R. A., M. Molina, J.E. Cox, L.J. Osher, and M.C. Piccolo. 2003. Stable carbon isotope ratio and composition of microbial fatty acids in tropical soils, Journal of Environmental Quality 32, 198–206.
Brye K.R., J.M. Norman, and S.T. Gower. 2002. Carbon budgets for a prairie and agroecosystems: effects of land use and interannual variability. Ecological Applications 12, 962–979.
Carney, K. M., B.A. Hungate, B.G. Drake, and J.P. Megonigal. 2007. Altered soil microbial community at elevated CO2 leads to loss of soil carbon. Proceedings of the National Academy of Sciences of the USA 104, 4990-4995.
Denef, K., J. Six, R. Merckx, and K. Paustian. 2004. Carbon sequestration in microaggregates of no-tillage soils with different clay mineralogy. Soil Science Society of America Journal 68, 1935–1944.
de Vries, F.T., J. Bloem, N. van Eekeren, L. Brussaard, and E. Hoffland. 2007. Fungal biomass in pastures increases with age and reduced N input. Soil Biology & Biochemistry 39, 1620-1630.
Elliott, E.T. 1986. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Science Society of America Journal 50, 627–633.
Elliott, E.T., R.V. Anderson, D.C. Coleman, and C.V. Cole. 1980. Habitable pore space and microbial trophic interactions. Oikos 35, 327–335.
Fargione, J.E., T.R. Cooper, D.J. Flaspohler, J. Hill, C. Lehman, T. McCoy, S. McLeod, E.J. Nelson, K.S. Oberhauser, and D. Tilman. 2009. Bioenergy and Wildlife: Threats and Opportunities for Grassland Conservation. Bioscience 59, 767-777.
Fierer, N., J.P. Schimel, and P.A. Holden. 2003. Variations in microbial community composition through two soil depth profiles. Soil Biology and Biochemistry 35, 167-176.
Follett, R.F., S.E. Samson-Liebig, J.M. Kimble, E.G. Preussner, and S.W. Waltman. 2001. Carbon sequestration under the CRP in the historic grassland soils of the USA. p. 27–49. In R. Lal et al (ed.) Soil management for enhancing carbon sequestration. Soil Science Society of America Spec. Publ. 57. SSSA, Madison, WI.
Food, Conservation, and Energy Act of 2008. Public Law 110-234.
Gebhart, D.L., H.B. Johnson, H.S. Mayeux, and H. W. Polley. 1994. The CRP increases soil organic carbon. Journal of Soil and Water Conservation 49, 488-492.
Guckert, J. B., M. A. Hood, and D. C. White. 1986. Phospholipid ester-linked fatty acid profile changes during nutrient deprivation of Vibrio cholerae: increases in the trans/cis ratio and proportions of cyclopropyl fatty acids. Applied and Environmental Microbiology 52, 794–801.
Hallett, P.D., D.S. Feeney, A.G. Bengough, M.C. Rillig, C.M. Scrimgeour, and I.M. Young. 2009. Disentangling the impact of AM fungi versus roots on soil structure and water transport. Plant and Soil 314, 183-196.
Harris, J. 2009. Soil Microbial Communities and Restoration Ecology: Facilitators or Followers? Science 325, 573-574.
Heggenstaller, A.H., K.J. Moore, M. Liebman, and R.P. Anex. 2009. Nitrogen Influences Biomass and Nutrient Partitioning by Perennial, Warm-Season Grasses. Agronomy Journal 101, 1363-1371.
Kaur, A., A. Chaudhary, A. Kaurl, R. Choudhary, and R. Kaushik. 2005. Phospholipid fatty acid – A bioindicator of environment monitoring and assessment in soil ecosystem. Current Science 89, 1103-1113.
Kieft T.L., D.B. Ringelberg, and D.C. White. 1994. Changes in Ester-Linked Phospholipid Fatty Acid Profiles of Subsurface Bacteria during Starvation and Desiccation in a Porous Medium. Applied and Environmental Microbiology 60, 3292–3299.
Kieft T.L., E. Wilch, K. O’Connor, D.B. Ringelberg, and D.C. White. 1997. Survival and phospholipid fatty acid profiles of surface and subsurface bacteria in natural sediment microcosms. Applied and Environmental Microbiology 63, 1531-1542.
Kucharik, C.J. 2007. Impact of prairie age and soil order on carbon and nitrogen sequestration. Soil Science Society of America Journal, 71: 430-441.
McKinley, V.L., A.D. Peacock, and D.C. White. 2005. Microbial community PLFA and PHB responses to ecosystem restoration in tallgrass prairie soils. Soil Biology & Biochemistry 37, 1946–1958.
Mentzer, J.L., R. Goodman, and T.C. Balser. 2006a. Linking soil process and microbial ecology in freshwater wetland ecosystems. Plant and Soil 289, 17-34.
Mentzer, J.L., R. Goodman, and T.C. Balser. 2006b. Microbial seasonal response to hydrologic and fertilization treatments in a simulated wet prairie. Plant and Soil 284, 85-100.
Oades, J.M. 1984. Soil organic matter and structural stability: mechanisms and implications for management. Plant and Soil 76, 319-337.
Oades J.M. and A.G. Waters. 1991. Aggregate hierarchy in soils. Australian Journal of Soil Research 29, 815-828.
Ratledge, C. and S.G. Wilkinson, S.G. 1988. Microbial Lipids. Academic Press, London.
Rillig, M.C., S.F. Wright, and V.T. Eviner. 2002. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: comparing effects of five plant species. Plant and Soil 238, 325–333.
Santruckova, H., M.I., Bird, Y.N. Kalaschnikov, M. Grund, D. Elhottova, M. Simek, S. Grigoryev, G. Gliexner, A. Arneth, and E.D. Schulze. 2003. Microbial characteristics of soils on a latitudinal transect in Siberia. Global Change Biology 9, 1106–1117.
Schutter M.E., and R.P. Dick. 2002. Microbial community profiles and activities among aggregates of winter fallow and cover-cropped soil. Soil Science Society of America Journal 66, 142–153.
Simpson R.T., S.D. Frey, J. Six, and R.K. Thiet. 2004. Preferential accumulation of microbial carbon in aggregate structures of no-tillage soils. Soil Science Society of America Journal 68, 1249–1255.
Six, J., S.D. Frey, R.K. Thiet, and K.M. Batten. 2006. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Science Society of America Journal 70, 555-569.
Smith, M. 2000. Land Retirement. In: Heimlich, R. (ed). Agriculture Handbook No. AH722: Agricultural Resources and Environmental Indicators (2003). U.S. Department of Agriculture, Washington, D.C. Available at: http://www.ers.usda.gov/publications/arei/ah722/dbgen.htm. Last accessed: Nov. 21, 2009.
Smithwick E.A.H., M.G. Turner, K.L. Metzger, and T.C. Balser. 2005. Variation in NH4+ mineralization and microbial communities with stand age in lodgepole pine (Pinus contorta) forests, Yellowstone National Park (USA). Soil Biology and Biochemistry 37, 1546-1559.
Staben, M.L., D.F. Bezdicek, J.L. Smith, and M.F. Fauci. 1997. Assessment of soil quality in Conservation Reserve Program and wheat-fallow soils. Soil Science Society of America Journal 61, 124-130.
Steenwerth, K.L., L.E. Jackson, F.J. Calderón, M.R. Stromberg, and K.M. Scow. 2003. Soil microbial community composition and land use history in cultivated and grassland ecosystems of coastal California. Soil Biology & Biochemistry 35, 489–500.
Thomas, G.W. 1996. Soil pH and soil acidity. In: Sparks, D.L. (ed.), Methods of Soil Analysis, Part 3. Chemical Methods. American Society of Agronomy, Inc.; Soil Science Society of America, Inc., Madison, WI, pp. 475-490.
U.S. Department of Agriculture. 2009a. “Agriculture Secretary Vilsack Announces $1.7 Billion in Conservation Reserve Program Rental Payments”. News Release No. 0497.09. Available at: http://www.usda.gov/wps/portal/!ut/p/_s.7_0_A/7_0_1OB ?contentidonly=true&contentid=2009/10/0497.xml. Last accessed: Oct. 26, 2009.
U.S. Department of Agriculture. 2009b. “CRP Enrollment as of September 2009 and October 2009 Rental Payments.” Available at: http://www.fsa.usda.gov/Internet/FSA_File/ apportstate.pdf. Last accessed: Oct. 26, 2009.
USDA Natural Resources Conservation Service (NRCS). 2007. National Resources Inventory 2003 Annual NRI: Land Use. Available at: http://www.nrcs.usda.gov/technical/NRI/2003/Landuse-mrb.pdf. Last accessed: Dec. 15, 2009.
U.S. Department of Energy (DOE). 2005. Biomass as feedstock for a bioenergy and bioproducts industry: the technical feasibility of billion-ton annual supply. 2005. Available at: http://www1.eere.energy.gov/biomass/pdfs/final_billionton_vision_report2.pdf. Last accessed: Dec. 17, 2009.
Vestal J.R. and D.C. White. 1989. Lipid analysis in microbial ecology: Quantitative approaches to the study of microbial communities. BioScience 39, 535-541.
White, D. C. 1993. In situ measurement of microbial biomass, community structure, and nutritional status. Philosophical Transactions of the Royal Society of London, Series 344A, 59-67.
White, D.C., W.M. Davis, J.S. Nickels, J.D. King, and R.J. Bobbie, R.J. 1979. Determination of the sedimentary microbial biomass by extractible lipid phosphate. Oecologia 40, 51–62.
Wilkinson S., J. Anderson, S. Scardelis, M. Tisiafouli, A. Taylor, and V. Wolters. 2002. PLFA profiles of microbial communities in decomposing conifer litters subject to moisture stress. Soil Biology and Biochemistry 34, 189–200.
Wilson, S.D., and D.C. Hartnett. 1997. Effects of mycorrhizae on plant growth and dynamics in experimental tallgrass prairie microcosms. American Journal of Botany 84, 478–482.
Zak, D.R., D.B. Ringelberg, K.S. Pregitzer, D.L. Randlett, D.C. White, and P.S. Curtis. 1996. Soil microbial communities beneath Populus grandidentata grown under elevated atmospheric CO2. Ecological Applications 6, 257–262.
Zelles, L. 1997. Phospholipid fatty acid profiles in selected members of soil microbial communities. Chemosphere 35, 275-294.
Zelles, L. 1999. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review. Biology and Fertility of Soils 29, 111–129.