Final Report for GNE10-001
Project Information
A field experiment conducted from 2007-2009 assessed the impact of various soil amendments on potato disease suppression and soil microbial community characteristics. To follow up on that experiment, this SARE project was designed to investigate the connection between specific pathogen levels for Rhizoctonia solani in soil and black scurf (caused by R. solani), as well as overall yields of potato. In order to achieve this goal, DNA was extracted from soil samples which had been collected in the summer and fall of each year of the field study. Quantitative polymerase chain reaction (qPCR) was then used to detect the amount of R. solani DNA in composite soil samples representing each treatment. Results of qPCR showed that R. solani was detected in almost all soil DNA samples from each season of each year. Levels of R. solani DNA were comparable (~10-15 g DNA or ~10 femtograms/g soil) among soils receiving different soil amendments. While there were many significant differences in the levels of R. solani DNA detected among treatments, these effects were often not observed in more than one year, or more than one season. Correlation analysis revealed that there was no overall correlation between the levels of R. solani DNA detected in the summer and fall samplings. However, there was an overall correlation between the levels of R. solani DNA in soils collected from the summer sampling and severity and incidence of black scurf on tubers, demonstrating that levels of R. solani DNA in the soil in the midst of the growing season are roughly indicative of the disease observed in the fall (at harvest). Of the treatments used in the study, compost amendment resulted in the most consistent trends from year to year, with increases in yield between 6% and 14%, and also slight increases in the severity (less than 0.5% increase in average overall surface area coverage) of black scurf disease in two of the three years.
Introduction:
Rhizoctonia solani is an important soil-borne pathogen of many crops, including wheat, rice, corn and potatoes. The species of R. solani is divided up into 13 anastamosis groups (AGs). Anastomosis refers to the fusion of hyphae from different strains of R. solani (strains which fuse belong to the same AG). R. solani affecting potatoes in Maine belongs largely to AG-3. R. solani causes two types of symptoms in potato; stem canker and black scurf of tubers. Stem canker is characterized by formation of lesions on the stems and stolons of potato plants that can grow to eventually girdle and kill the plant if left unchecked. Black scurf of tubers represents the survival structures of the pathogen and therefore provides a future source of inoculum in seed tubers, as well as affecting marketability of table stock. Soil conditions in Maine and other parts of New England, because of their cool and wet climate during the growing season, are favorable to the fungus and rhizoctonia disease development.
Growers often struggle with ineffective chemical controls for rhizoctonia disease, such as seed treatments, which are also economically and environmentally costly. Several biological control organisms including the bacterium Bacillus subtilis, the fungus Trichoderma virens and a non disease-causing isolate of R. solani AG-3 (Rhs1A1) have all been shown to control rhizoctonia disease in various plant species including potato. For example, antibiotics produced by B. subtilis have been shown to work against R. solani (Asaka and Shoda, 1996), and amendment of soils with T. virens is known to suppress R. solani populations via mycoparasitism (Harman, 2006; Hoitinck and Boehm, 1999). A novel hypovirulent of R. solani (designated as Rhs 1A1) has also been shown to suppress black scurf and stem canker and promote plant growth. This is due to decreased levels of phenylacetic acid (PAA), which is phytotoxic at high levels but acts as a plant growth-promoting hormone in low physiological doses (Bandy and Tavantzis, 1990; Jian et al., 1997; Liu et al., 2003).
Compost amendments have been shown to suppress rhizoctonia diseases by serving as a substrate for growth of beneficial microorganisms like T. virens (Hoitinck and Boehm, 1999). Composts rich in pine barks also have the potential to control rhizoctonia disease (Hoitinck and Boehm, 1999, 12). As they decay, pine barks release quinic acid, which has been shown to also induce a state of hypovirulence in normal, virulent isolates of R. solani (Liu et al., 2003). Similarly, Brassica green manures are known to suppress some plant diseases through the process of biofumigation, when decomposing plant tissues release antimicrobial secondary metabolites (Larkin and Griffin, 2007).
Several of these treatments were recently used in a field trial to assess their efficacy at controlling rhizoctonia disease of potato and increasing yield. Results obtained from this recent field work have presented a complex picture regarding the role of these soil amendments in controlling rhizoctonia disease of potato, and improving soil quality and crop yields. Soil microbial community characteristics, including overall bacterial and fungal counts and general microbial activity exhibited variable responses to different soil amendments (Bernard et al., unpublished data). Disease and yield data also varied depending on treatment or combination of treatments (Bernard et al., unpublished data). The methods used in these experiments focused on broad differences in order to discern any basic trends. However, given the complexity of the information generated from the recent field project, it is crucial that we ask a specific question about how the biological controls, compost, and different rotation crops directly affect the soil-borne R. solani population.
The objectives of this study were to:
1. Monitor Rhizoctonia solani levels in the soils over the course of the growing season in response to the amendment of soils with a conifer-based compost, a rapeseed rotation crop, and the biological control fungi, hypovirulent Rhizoctonia solani (Rhs1A1) and Trichoderma virens, and the biological control bacterium Bacillus subtilis.
2. Correlate Rhizoctonia solani levels in the soil with observed tuber diseases and tuber yield data.
Both objectives of the study were successfully completed. Quantitative real-time PCR (qPCR) data were collected and analyzed for differences in levels of R. solani in amended soils from all three years of the original field study. These data were then correlated with disease and yield data from the same field study in order to determine the relationship between levels of R. solani DNA in soils with the levels of black scurf and the impact on tuber yield.
Cooperators
Research
In the spring of 2007, research plots were planted with potato at Aroostook Research Farm, Presque Isle, Maine. Plots were arranged in a randomized split block design (four replications), with half of each block receiving a compost amendment (Cobscook blend, comprised of hemlock bark, salmon, blueberry, cow manure, sphagnum peat, and limestone and produced by Coast of Maine Organic Products, Inc., Portland, ME). Plots in each block received one of three biocontrol agents or were left unamended. The biocontrol agents employed were Bacillus subtilis (Kodiak®, Bayer CropScience LP, Research Triangle Park, NC), Trichoderma virens (SoilGard®, Olympic Horticultural Products, Mainland, PA), and a hypovirulent strain of Rhizoctonia solani (Rhs1A1, Bandy and Tavantzis, 1990). Research plots consisted of six rows (each 7.62 meters long) with the four middle rows receiving amendment and the two outer rows serving as buffers between plots. Soil samples were collected at three times over the course of the growing season: prior to planting, two weeks post-emergence, and two weeks after harvest. Samples were collected in 8 cores from the two central rows of each plot to a depth of 20 cm. Soil samples were sieved to remove rocks and plant materials and stored at 4° C. The soil samples were then analyzed for different aspects of microbial ecology, including total bacterial and fungal counts, as well as assessment for community-level diversity among amendments. Following harvest, 50 tubers from each plot were also assessed for a variety of plant diseases, including black scurf caused by Rhizoctonia solani. Subsequently, in 2008 and 2009, plots were re-established at the same agricultural sites and a Brassica napus rotation was included, such that one half of the field plots followed rotation with a Brassica napus cover crop, and half of the plots followed a standard barley rotation. Soil samples were again collected in the same manner and similar analyses were done for microbial community profiles, tuber diseases and yield.
DNA was extracted from soil samples from each plot using the Fisher BioReagents SurePrep Soil DNA Isolation Kit (Fisher BioReagents, Fair Lawn, NJ), according to the manufacturer’s instructions. Extracted DNA was then combined by treatment and purified using the QIAquick PCR Purification Kit (Qiagen, Valencia, CA), according to the manufacturer’s instructions. Extracted DNA was quantitated using the Qubit™ Quantitation Platform (Invitrogen Corporation, Carlsbad, CA), before proceeding with qPCR. The extracted DNA was diluted to the same starting concentration of DNA within each year before quantitative PCR. Primers RsTqF1 and RsTqR1 used for quantitative PCR were described by Lees et al. (2003). These primers were tested on bulk soil DNA from control and amended plots as well as R. solani AG-3 genomic DNA and products were sequenced to confirm their identity. Quantitative PCR was performed using iCycler iQ™ 96 well PCR plates using the MyiQ™ Single-Color Real-Time PCR Detection System (Life Science Research, Hercules, CA). The 20 ?L reaction mix included iQ™ SYBR® Green SuperMix (Life Science Research, Hercules, CA), primers RsTqF1 and RsTqF2 at a final concentration of 0.2 ?M, and 3 ?L of template DNA. Starting quantities of the target sequence were calculated using a standard curve of Ct values generated from known amounts of R. solani AG-3 genomic DNA (from Rhs1A1).
Duncan’s Multiple Range Test at p=0.05 was used for mean separation of qPCR data for planned comparisons. Correlation of qPCR data with disease and yield data was achieved using Pearson’s correlation coefficient. All statistical analyses were carried out using Systems Analysis Software (SAS) Version 9.2 (SAS Institute Inc., Cary, North Carolina).
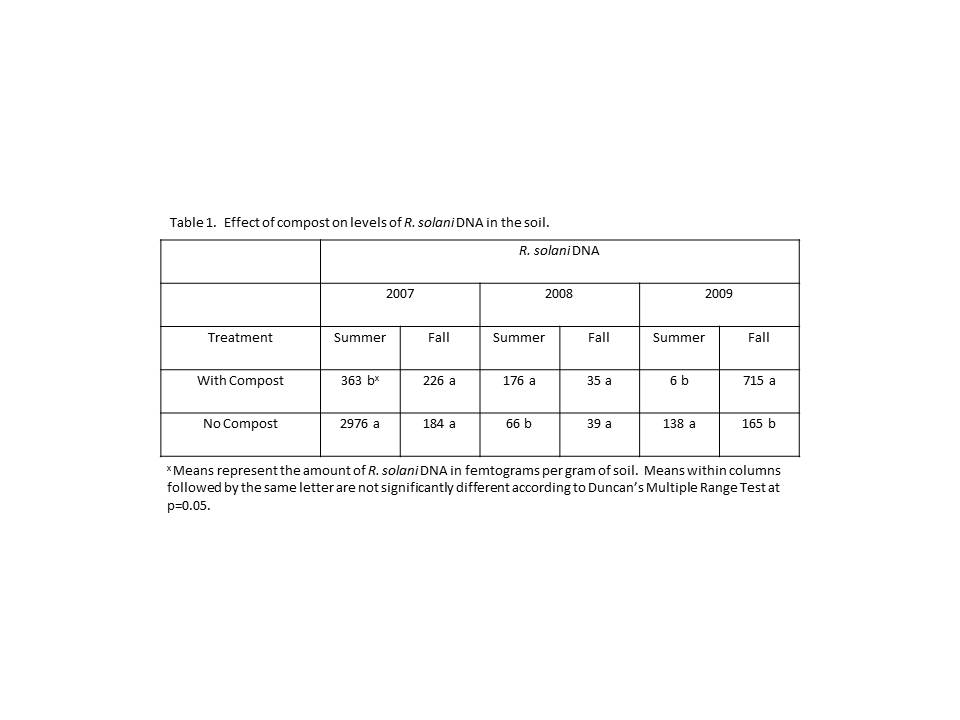
Analysis of the data collected in this study revealed few overall trends. Levels of R. solani in amended soils varied greatly from year-to-year. Compost amendment had significantly different effects in different years (Table 1). In 2007, levels of R. solani were significantly higher in non-composted plots (2976 femtograms of DNA) when compared to composted plots (363 femtograms) in the summer, while no difference was observed in the fall. In 2008, levels of R. solani DNA were significantly higher in composted plots (176 fg) when compared to non-composted plots (66 fg) in the summer, but were again not different in the fall. R. solani DNA levels were again higher in non-composted plots (138 fg) in 2009 than in composted plots (6 fg) in the summer, while the reverse was observed in the fall with higher levels in composted plots (715 fg) when contrasted with non-composted plots (165 fg).
Total yield measurements from all three years were significantly higher in plots amended with compost when compared to plots lacking compost. The increase in yield ranged from 6% in 2008 to approximately 14% in 2009 (the increase in 2007 was ~12%). Compost amendment, however, resulted in significant increases in the incidence of black scurf in both 2008 and 2009. The increase in incidence was approximately 24% in 2008 and 12% in 2009. Similar results were observed in the severity of black scurf in those years. Black scurf severity was 64% higher in composted plots than non-composted plots in 2008 and 16% higher in 2009. In 2007, black scurf severity was significantly lower in compost amended plots by about 13% when compared with non-composted plots. Incidence was not statistically different between composted and non-composted plots in 2007.
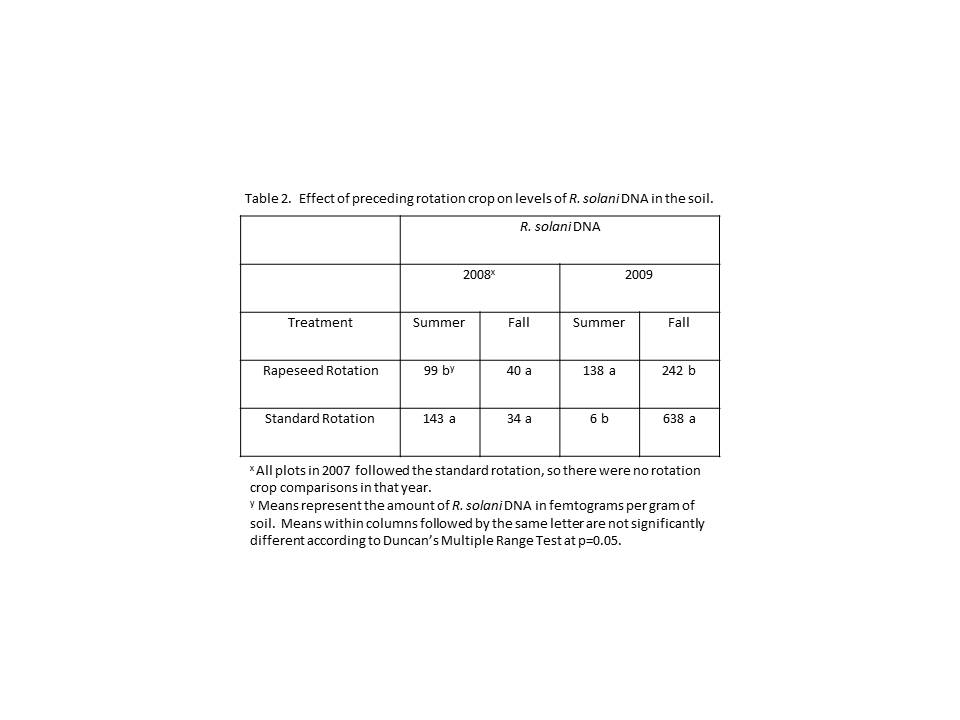
Rapeseed rotation had significant effects on levels of R. solani in the soil in both years that it was incorporated. However, the effects seen were different in both seasons of both years (Table 2). In 2008, summer levels of R. solani DNA were higher in plots following the standard rotation (143 fg) than those following rapeseed (99 fg), while in the fall the difference was not significant. In 2009, significantly higher levels of R. solani were found in plots following rapeseed rotation (138 fg) than the standard rotation (6 fg) in the summer, while in the fall plots following rapeseed rotation (242 fg) had significantly less R. solani DNA than plots that were under the standard rotation (638 fg).
Total yield and marketable yields were not significantly different between plots following rapeseed rotation when compared with those following the standard rotation. Severity of black scurf was not different between plots which followed the standard rotation and those which followed rapeseed. In one year of the study (2008), the incidence of black scurf was 9% lower in plots following rapeseed when compared to plots following the standard rotation.
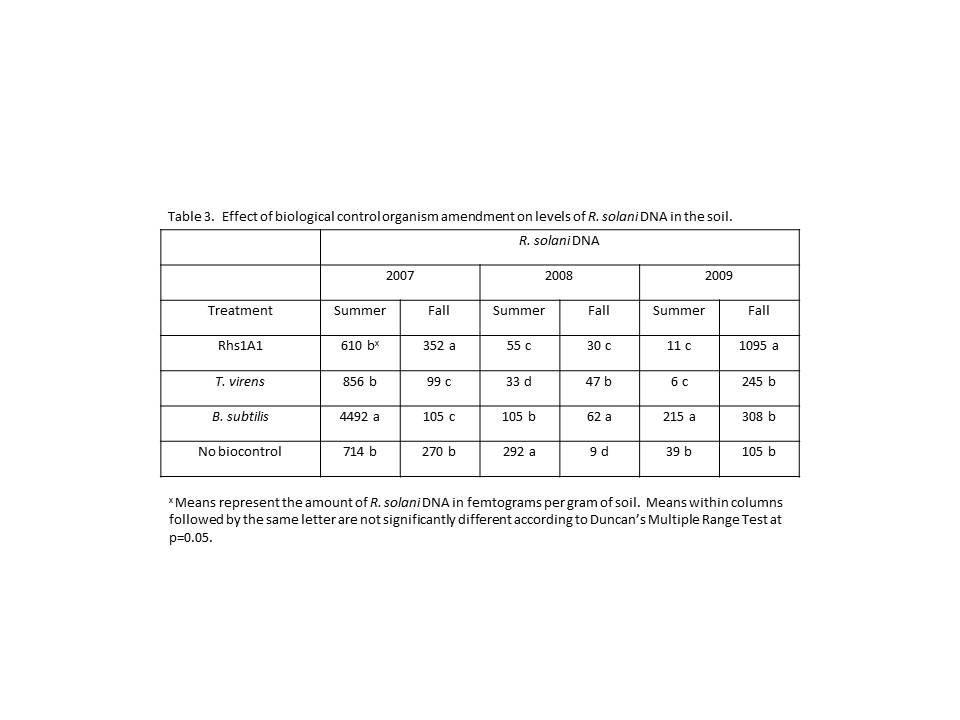
Biocontrol amendments yielded the most consistent results between years and seasons (Table 3). In 2007, plots amended with B. subtilis had significantly higher levels of R. solani than all other plots in the summer soils, while in the fall plots amended with Rhs1A1 (352 fg) had significantly higher levels of R. solani DNA than those in untreated plots (270 fg), while B. subtilis (105 fg) and T. virens (99 fg) amended plots had less than the untreated plots. Levels of R. solani DNA were significantly lower in 2008 in Rhs1A1 (55 fg), B. subtilis (105 fg), and T. virens (33 fg) than untreated plots (292 fg) in the summer, while all plots receiving a biocontrol organism were higher in levels of R. solani DNA than untreated plots in the fall soils. In the final year of the study, levels of R. solani DNA were higher in plots receiving B. subtilis (215 fg) than untreated plots (39 fg), which had higher levels than plots receiving Rhs1A1 (11 fg) and T. virens (6 fg) in the summer. In the fall of the same year, R. solani DNA was higher in plots receiving Rhs1A1 (1095 fg) than all other plots (105-308 fg).
Biocontrol amendment only affected yields in 2008, when plots amended with Rhs1A1 showed a significant decrease in yield compared to plots not receiving any biocontrol amendment. Biocontrol amendment only affected black scurf incidence and severity in a single year (2007) of the study. In that year, both incidence and severity were significantly lower in plots amended with Rhs1A1 when compared plots not receiving a biological control amendment. The reduction in severity was approximately 23%, while the reduction in incidence was about 13%.
There was an overall correlation between the levels of R. solani DNA in the soil in the summer with the severity (r=0.238, p=0.033) and incidence (r=0.29146, p=0.009) of black scurf. The incidence and severity of black scurf were found not to be correlated with yield, either. The levels of R. solani DNA in the fall were not correlated with anything when data from plots amended with Rhs1A1 were included. When plots receiving Rhs1A1 were excluded, fall qPCR levels were correlated with overall tuber yield (r=0.31933, p=0.0129). There was no correlation between the summer qPCR data and the fall qPCR data under any circumstances.
The overall aims of this study were to determine the impact of 1) pathogen levels in the soil on tuber disease and yield and 2) particular disease-suppressive treatments on pathogen levels in the soil. The results of this study were somewhat unexpected, as the levels of R. solani in the soil did not always correlate with the amount of disease that was seen. For instance, compost amended plots had significantly higher black scurf severity in 2009, but the opposite was true for levels of R. solani in the summer. Similarly, increased levels of R. solani in plots amended with the biocontrol bacterium B. subtilis did not show corresponding increases or decreases in the severity or incidence of black scurf. This indicates that levels of the pathogen are not necessarily the most important factor governing the development of black scurf. Also the observed increases in black scurf severity represent an average tuber surface area coverage of less than 1.5%, which would not be large enough to result in increased economic loss due to black scurf.
Another possible explanation for the lack of correlation between levels of pathogen in the soil and development of black scurf observed in this study may be related to the lack of a selective method for assaying only pathogenic isolates of R. solani AG-3. While R. solani AG-3 is the major causative agent of black scurf in Maine, not all isolates have the same-disease causing capability, and some may in fact be mutualistic(i.e. Rhs1A1). Since the assay used in this study was specific for AG-3 it was only capable of determining the levels of the entire AG-3 community in the soil, and not only pathogenic isolates. However, higher levels of R. solani DNA were expected in plots amended with Rhs1A1 (since Rhs1A1 would have also been detected with qPCR) than plots not receiving a biocontrol, which was not always the case. This demonstrates that added Rhs1A1 represent a small, insignificant proportion of the R. solani soil population in the soil or that the Rhs1A1 is not surviving in the soil until emergence of the plant. In contrast, this data could indicate that the natural population of R. solani survives better when not in competition with the hypovirulent isolate.
Asaka, O., and Shoda, M. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Appl. Environ. Microbiol. 1996. 62: 397-404.
Bandy, B.P., and Tavantzis, S.M. Effect of Hypovirulent Rhizoctonia solani on Rhizoctonia Disease, Growth, and Development of Potato Plants. Am Potato J. 1990. 189-199.
Harman, G. E. Overview of mechanisms and Uses of Trichoderma spp. Phytopath. 2006. 96: 190-194.
Hoitinck, H.A.J., and Boehm, M.J. Biocontrol within the Context of Soil Microbial Communities: A Substrate-Dependent Phenomenon. Ann. Rev. Phytopath. 1999. 37: 427-446.
Jian, J., Lakshman, D.K., and Tavantzis, S.M. Association of Distinct Double-Stranded RNAs with Enhanced or Diminished Virulence in Rhizoctonia solani Infecting Potato. Mol. Plant Micro. Inter. 1997. 10: 1002-1009.
Larkin, R.P., and Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop Protect. 2007. 26: 1067-1077.
Lees, A.K., Cullen, D.W., Sullivan, L., and Nicolson, M.J. Development of conventional and quantitative real-time PCR assays for the detection and identification of Rhizoctonia solani AG-3 in potato and soil. Plant Path. 2002. 51: 293-302.
Liu, C., Lakshman, D.K., and Tavantzis, S. Quinic acid induces hypovirulence and expression of a hypovirulence-associated double-stranded RNA in Rhizoctonia solani. Curr. Genet. 2003. 43: 103-111.
{kind=link}
{kind=link}
{kind=link}
There are several important outcomes to this research. First and foremost, the collection of quantitative data from field soil samples about an important pathogen such as R. solani is an important facet of assessing management strategies. While some studies have used quantitative PCR as a diagnostic tool for soil-borne pathogens, few, if any, have used it to assess the direct efficacy of soil amendments. Fewer still have used this data to investigate impacts on yield and disease. In this study both of these issues were addressed.
The quantitative data collected showed an overall positive correlation between summer qPCR data and incidence and severity of black scurf, confirming that controlling levels of R. solani in the soil has an important place in a long-term management strategy. The lack of correlation between fall qPCR data and severity and incidence of black scurf is also an important result further demonstrating that the presence and level of the pathogen in the soil at the time of tuber harvest is not as important in gauging the severity of black scurf as earlier in the season as weather conditions might be.
Education & outreach activities and participation summary
Participation summary:
While no outreach has been achieved to this point in the project, due to the extensive amount of data collected and analyzed, several plans are in place for outreach in the near future. The original research project that included the field trials used in this study was set up with a Cooperative Extension agent (John Jemison) serving on the team. Additionally, research talks are planned for the Annual Maine Agricultural Trades Show in January 2012, and the Northeast Potato Technology Farm in March 2012. A manuscript is also being prepared for submission to an appropriate journal this fall.
Project Outcomes
This study has shown that it is possible to detect R. solani in a modern agricultural setting, and to utilize that data to answer a variety of ecological, biological, and practical questions. Assuming that a facility is equipped with the necessary tools for qPCR it is possible to determine the material cost per soil sample for detection of R. solani. A rough estimate of this material cost is $17.50 per soil sample. This estimate only includes the cost of materials specifically required for qPCR and excludes the cost of labor. Labor costs are not easy to estimate as a single sample would not be prepared for qPCR by itself (as qPCR is carried out on 96-well plates). In fact, it is possible to process up to 24 soil samples at a time given the limitation of required hardware.
Farmer Adoption
It is quite difficult to predict the reaction of the growers to our data as we are still in the process of distilling our results in terms of prioritizing the significance of the different aspects of our findings.
Areas needing additional study
While this study revealed much about the response of R. solani to various soil amendments, much more temporal data should be collected. A single date for sampling in the summer and the fall limits our understanding as to what is going on throughout the growing season. Additional sampling dates in future field trials in conjunction with fewer treatments would provide data on how the pathogen behaves in response to particular treatments at different stages of plant growth and may provide even more accurate predictors of tuber disease levels. Similarly, residual effects from the previous year’s amendments were not addressed in this study as the research plots were moved to neighboring tracts of land each year. Additionally, finding a practical way of monitoring not only the pathogen in the soil, but also the biological control organisms or known groups of soil antagonists would shed more light on the ecological impacts of soil amendments such as biocontrol organisms and composts.