Final report for GNE16-121
Project Information
Swede midge, an invasive pest of Brassica vegetable crops, is currently causing up to 100% yield loss of organic broccoli and kale in the Northeast. Swede midge larvae feed within the new leaves of the plant growing point and are protected from contact with foliar insecticides. Only one swede midge larva can render a heading Brassica plant unmarketable. This insect is particularly difficult for small-scale organic growers because they lack adequate space to rotate away from land infested with swede midge, and because there are no effective insecticides for swede midge management that are allowable for organic production. Pheromone mating disruption, which involves confusing males with unnaturally-high doses of synthetic sex pheromone, is particularly promising for swede midge management for three reasons: 1) Pheromones are highly species-specific, limiting impacts on non-target organisms; 2) It is allowable for organic production; and 3) It prevents mating and subsequent larval feeding within the crop. However, using a synthetic blend of the same compounds naturally produced by female midges is too expensive. Cheaper, unpurified structural variants, "racemic" blends, and reduced-component blends are cheaper, but their efficacy for mating disruption is currently unknown. Additionally, effects of unnatural pheromones on male and female behaviors are not well understood. Our research explored the effects of mating disruption treatments on male and female mate-seeking behaviors, as well as tested economical racemic and single-component mating disruption systems in Brassica crops in the field.
Despite the potential for chiral and racemic single-component pheromone blends to reduce the costs of swede midge mating disruption, we found that they were ineffective in confusing males and providing broccoli crop protection. Based on our results from these projects and a previous (USDA NIFA Crop Protection and Pest Management) grant, we have determined that the three-component chiral pheromone blend, which is the blend naturally-produced by females, is the most effective for swede midge mating disruption. We found that females were less likely to mate following three-component chiral pheromone pre-exposure, and that males were less likely to locate female mimics after chiral pheromone pre-exposure. In our USDA grant field trials, the three-component chiral blend was the only blend to both confuse males and provide significant crop protection in at least one out of the two years of our trials. Therefore, we believe that this is the most promising blend for mating disruption of swede midge. Additional research will be necessary to test alternative ways of reducing the cost of swede midge mating disruption, such as through the use of other dispenser types and densities.
We believe that we are the first to document "autodetection," the ability of females to detect and respond to their own pheromone, in the family of galling flies, Cecidomyidae. Females are typically overlooked in mating disruption, and our study adds to the growing body of literature that supports the idea that females can alter their behavior in important ways that can influence the success of mating disruption.
Pheromone mating disruption, although highly successful for moth pests in perennial crops, has not been as widely adopted for managing non-lepidopteran pests, particularly those in annual cropping systems. In order to use mating disruption for flies in vegetable systems, a thorough understanding of how they respond to pheromones will be necessary. Flies may respond to pheromones differently than moths. Additionally, migration patterns of pests within annual crop rotations is an issue that needs to be addressed for mating disruption to function effectively in vegetable cropping systems. For ecologically-based pest management to advance to improve agricultural sustainability, continuing research is needed to fully understand the biology and ecology of insect pests. Effective and economical pheromone mating disruption systems would provide vegetable growers with another tool for managing challenging pests. However, for swede midge, more research is necessary.
We aimed to test lower-cost pheromone blends for mating disruption of swede midge, an invasive fly that attacks Brassica vegetables, and to better understand how synthetic pheromones influence midge behavior through the following objectives:
Objective 1. Test the ability of pheromone blends to elicit non-competitive mechanisms of disruption.
How pheromone mating disruption serves to disrupt mating is categorized into two types of mechanisms, competitive and non-competitive1. Under competitive disruption, females compete for attention by males with pheromone dispensers, which attract the males away from the females. By contrast, under non-competitive disruption, pheromone plumes emitted by dispensers are unattractive and render the males incapable of responding to either females or dispensers. Under competitive disruption, efficacy can decline with increasing insect populations. Non-competitive disruption mechanisms, including desensitization of males and adverse effects on females, can enhance the efficacy of the system even when populations are large. Knowing which mechanisms are at play with given pheromone mating disruption systems can inform decisions relating to pheromone blend, dispenser type, and dispenser densities to enhance the efficacy of the system.
We aimed to test the effects of natural (chiral), racemic, and single-component pheromone blends (the "main" component only) on both male and female behavior using laboratory and field trials to inform us of the mechanisms responsible for mating disruption. Pheromone blends that both impair male ability to locate mates and reduce female receptivity to males will be the best candidates for swede midge mating disruption.
Objective 1.1. Compare the abilities of pheromone blends to desensitize males and to impair their ability to locate females.
We pre-exposed males to field-relevant, high doses of pheromones, and then observed their ability to find females in a y-tube olfactometer. Blends that desensitize males and prevent them from locating mates will be more effective in a pheromone mating disruption system.

Objective 1.2. Test the effect of large doses of pheromones on the reproductive behavior of female midges.
To test whether large doses of pheromones influenced female reproductive behavior, we observed female pheromone-releasing behavior, or “calling,” and their propensity to mate during and following pheromone pre-exposure in flasks with different pheromone blend treatments (Fig. 1). Pheromone blends that delay or reduce calling and render females unreceptive to males will enhance the efficacy of a mating disruption system.
Objective 2. Test the effect of swede midge population density on the efficacy of pheromone blends for mating disruption in enclosures.
We compared the abilities of chiral and racemic three-component pheromone blends to disrupt mating and reduce numbers of feeding larvae on plants in enclosures set up in the field containing varying numbers of midges.

Objective 3. Conduct open-field trials of one-component blends to test their ability to disrupt mating and provide crop protection.
Using broccoli as a test crop, we installed dispensers in test plots releasing either the chiral major pheromone component or the racemic major pheromone component (Fig. 2). We tested whether the pheromone treatments successfully disrupted mating and provided crop protection using trap shutdown, crop damage assessment and broccoli yield measurements.

Swede midge (Contarinia nasturtii; Diptera: Cecidomyiidae), a small galling fly, is an invasive insect pest of Brassica vegetable crops that is threatening the livelihood of organic vegetable growers in the Northeast (Fig. 3). Adult flies lay their eggs into the growing points of their host crops, including broccoli, kale, and cabbage, and larval feeding causes distorted growth of the plant (Fig. 4).

There are currently no effective, economically-feasible organic management strategies to defend against crop damage from swede midge. Growers in effected areas in the Northeastern U.S. are experiencing devastating crop loss, to the point that some are considering abandonment of Brassica vegetable production entirely. Others are turning toward use of neonicotinoid insecticides followed by calendar sprays of conventional foliar insecticides to protect conventionally grown crops.
Pheromone mating disruption is a promising tactic that has been very successful for pests of other horticultural crops and involves the release of large quantities of insect sex pheromones to confuse males so that they cannot locate their mates. Swede midge females release a blend of three chiral pheromone components (2S,9S-diacetoxyundecane, 2S,10S-diacetoxyundecane, and 2S-acetoxyundecane in a 1:2:0.02 ratio) to attract males for mating. This natural, attractive pheromone blend for swede midge is too costly to be commercially feasible for mating disruption. Cheaper, chemically similar, repellent pheromone variants ("racemic" blends, Fig. 5 and Table 1)

are available for swede midge, but their efficacy for pheromone mating disruption is unknown. Eliminating synthesis of two of the swede midge pheromone components and using only the "main" component for mating disruption would allow for considerable cost savings. This project challenged the assumption that the most attractive pheromone blend for an insect is the most effective for mating disruption, and compared the behavioral effects of the natural (chiral) blend with lower cost, repellent blends. Little is known about how racemic and single-component blends function for swede midge mating disruption, and even less is known about how female flies respond to synthetic pheromones.
Table 1. Swede midge natural pheromone compounds and compounds produced in the synthetic racemic blend. (2S,10S)-diacetoxyundecane is the “main” pheromone component in the pheromone blend produced by females. Compounds in the racemic blend are produced during laboratory synthesis of the swede midge pheromone and are cheaper to produce than the naturally-produced compounds.
|
Ratio in blend2 |
Naturally-produced compound |
Compounds in synthetic racemic blend |
Attractiveness of racemic compounds3 |
|
|
1 |
(2S,9S)- diacetoxyundecane |
(2S,9R)- (2S,9S)- (2R,9S)- (2R,9R)- diacetoxyundecane 1:1:1:1 ratio |
Attractive |
|
|
2 |
(2S,10S)- diacetoxyundecane |
(2S,10S)- (2S,10R)- meso-(2,10) diacetoxyundecane 1:1:2 ratio |
Not attractive (repellent) |
|
|
0.02 |
(2S)- acetoxyundecane |
(2S)- (2R)- acetoxyundecane 1:1 ratio |
Attractive |
|
Cooperators
- (Researcher)
Research
Objective 1. Test the ability of pheromone blends to elicit non-competitive mechanisms of disruption.
Experiment 1.1. Male swede midge mate-seeking behavior following pheromone pre-exposure.
For some insect species, pre-exposure to large doses of pheromone desensitizes males and renders them incapable of responding normally to pheromones afterward. Synthetic pheromone blends that prevent males from responding normally to pheromone signals released by females can enhance the efficacy of mating disruption. In the laboratory, we pre-exposed male midges to either a low or high dose of three-component chiral or racemic pheromones (Table 2) for ten minutes, and then placed them into a y-tube olfactometer. A y-tube olfactometer is a device that allows an insect to choose between two different odor sources. We gave the males a choice between a solvent-only control and a female-mimicking attractive pheromone dose. We tested whether pheromone blend and dose influenced the likelihood that males were able to locate the female mimic using a binary logistic regression.
Table 2. Pheromone treatments used in male desensitization experimentz
|
Pheromone blend |
Diacetoxyundecane |
Acetoxyundecane |
||||
|
2S,9S |
2,9 |
2S,10S |
2,10 |
2,S |
2 |
|
|
Three-component chiral (High dose) |
10 ng |
- |
20 ng |
- |
0.2 ng |
- |
|
Three-component chiral (Low dose) |
1 ng |
- |
2 ng |
- |
0.02 ng |
- |
|
Three-component racemic (High dose) |
- |
40 ng |
- |
80 ng |
- |
0.4 ng |
|
Three-component racemic (low dose) |
- |
4 ng |
- |
8 ng |
- |
0.04 ng |
|
Control (solvent only) |
- |
- |
- |
- |
- |
- |
zThe natural S stereoisomers comprise either one quarter or one half of each racemic blend, and thus four times or twice as much racemic material was needed to deliver the same amounts of each S stereoisomer as in the chiral blend2.
Experiment 1.2. Effect of pheromone exposure on reproductive behavior of females.
To date, few studies have documented whether or not female insects are capable of "autodetecting" and responding to pheromone mating disruption treatments. In some moth species, females increase the frequency of their own pheromone-releasing behavior and are less likely to mate after pheromone pre-exposure4. The objectives of these experiments were to determine whether a) swede midge females increase "calling" (pheromone-releasing) behavior when exposed to synthetic pheromone; and b) female propensity to mate changes following pheromone pre-exposure.
Calling
For the first experiment, we took newly emerged, virgin females from a wild Ontario population and placed them singly into flasks containing pheromone wicks (Fig. 6) and observed their calling behavior in the laboratory for four hours. We first conducted a dose-response experiment to determine which pheromone dosages elicited responses from females using two doses of the three-component chiral blend (Table 3). We chose these doses because they are high enough to confuse males in a laboratory (pers. obs.). Because we found no differences in female calling behavior between low and high doses, we used the low dose for our blend experiment. In the blend experiment, the wicks released either the three-component chiral pheromone blend, the three-component racemic blend, the chiral main pheromone component, the racemic main pheromone component, or a solvent-only control (Table 3). Air circulated through the flasks using tubing connected to a small air compressor, and pheromone-laden air leaving the flasks was removed from the room via an overhead exhaust duct. We checked the females every fifteen minutes from 9:30 am through 1:30 pm, their peak period of calling, and recorded whether or not they were releasing pheromones by checking to see if their ovipositors and pheromone glands were extruded.
Table 3. Pheromone blends used in autodetection experiments
|
Pheromone blend |
Diacetoxyundecane |
Acetoxyundecane |
||||
|
2S,9S |
2,9 |
2S,10S |
2,10 |
2,S |
2 |
|
|
Dose-response experiment |
||||||
|
Three-component chiral (High dose) |
10 ng |
- |
20 ng |
- |
0.2 ng |
- |
|
Three-component chiral (Low dose) |
1 ng |
- |
2 ng |
- |
0.02 ng |
- |
|
Control (solvent only) |
- |
- |
- |
- |
- |
- |
|
Blend-response experiment |
||||||
|
Three-component chiral (3C-N) |
1 ng |
- |
2 ng |
- |
0.02 ng |
- |
|
Three-component racemic (3C-R) |
- |
4 ng |
- |
8 ng |
- |
0.04 ng |
|
Single-component chiral (1C-N) |
- |
- |
2 ng |
- |
- |
- |
|
Single-component racemic (1C-R) |
- |
- |
- |
8 ng |
- |
- |
|
Control (solvent only) |
- |
- |
- |
- |
- |
- |
We repeated the calling experiment in 52x52 ft plots of broccoli treated with mating disruption to better understand how females react to synthetic pheromone in realistic field settings. The plots were set up at the University of Guelph Elora Research Station in Elora, Ontario. Each plot contained 20 rows of 'Everest' broccoli with 80 mating disruption dispensers. We used reservoir-type semi-permeable polyethylene bag dispensers. Each bag contained a microcentrifuge tube with a small hole drilled in the top, containing 50 ug (2S,9S)-diacetoxyundecane, 100 ug (2S,10S)-diacetoxyundecane, and 1 ug (2S)-acetoxyundecane (the three-component chiral blend). The bags hung from wire stakes 25 cm from the ground.
We held newly-emerged unmated females individually in small glass vials with mesh covers to allow for air circulation, and placed half of the vials in untreated plots and half in plots treated with three-component chiral pheromone mating disruption dispensers. Vials of midges sat atop overturned pots so that they were approximately 25 cm above the ground at the same height as the dispensers. We observed the females in both treated and untreated plots concurrently from 9:30 am until 12:30 pm and recorded whether or not they were releasing pheromones every ten minutes. Although we were only able to observe female calling behavior in one treated and one untreated plot at a time, we repeated the experiment across six experimental periods and two plots from each treatment in late July and August in 2017 and 2018, during 6 – 12 weeks after installing dispensers. During each period, we observed the calling behavior of 4-10 midges.
To test the effect of pheromone mating disruption treatments on the probability of female calling in our laboratory and field experiments, we used generalized linear mixed models (GLM) with generalized estimating equations (GEE) extensions. Calling followed a binomial distribution. We also tested whether pheromone treatments influenced the duration of calling. We used a binary scoring system (yes/no) to record whether we observed midges calling for at least half of the time period, and analyzed the effects of pheromone treatments on the probability of midges "continuously calling" using a binary logistic regression.
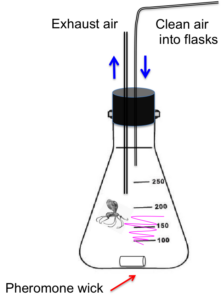
Propensity to mate
In the second part of our experiments, we took newly emerged, virgin females from our Swiss laboratory colony reared at the University of Vermont, pre-exposed them three-component chiral and racemic pheromone blends in flasks using the same dosages as in our blend-response calling experiments (Table 3, Fig 6), and then observed their propensity to mate with males. We omitted the single-component pheromone treatments from these experiments because they did not elicit any behavioral changes in the females in the first experiment (results detailed below). Using the same flask setup as the first experiment, we exposed the females to pheromone treatments for 10 minutes, removed them from the flasks, and then housed them in small containers with four males. We then observed the following behaviors: 1) Female calling every two minutes; 2) Female rejection of males following male courtship behavior; and 3) Successful copulation.
We used a binary logistic regression to determine whether pheromone treatments significantly influenced female mating success following pre-exposure. Similarly to our calling experiments, we used a GLM model with GEE extension to determine whether the probability of calling differed over time between pheromone treatments for midges that did not mate. Finally, we used another binary logistic regression model to determine whether the probability of females rejecting courting males differed between pheromone treatments.
Objective 2. Test the effect of swede midge population density on the efficacy of pheromone blends for mating disruption in enclosures.
Experiment 2. Population density effects on pheromone mating disruption efficacy.
We tested the efficacy of three-component chiral and racemic pheromone blends to disrupt mating using two densities of swede midge held in cages with cauliflower plants. Two population densities of newly emerged, unmated midges, either 2 females with 3 males, or 10 females with 15 males, were held in each cage. Each cage contained four potted cauliflower plants and received either a dispenser releasing the three-component chiral blend (50 ug (2S,9S)-diacetoxyundecane, 100 ug (2S,10S)-diacetoxyundecane, and 1 ug (2S)-acetoxyundecane), the three-component racemic blend (200 ug 2,9-diacetoxyundecane, 400 ug 2,10-diacetoxyundecane, and 2 ug 2-acetoxyundecane) or no dispenser (control). We situated the cages at least 50 m apart and separated them by buildings to prevent the spread of pheromone treatments between cages. After allowing the midges three days to mate, we returned them to the laboratory, where we removed the plants and placed them in separate midge-free cages to allow the eggs to develop into larvae. After 10 days, we dissected the growing points of each plant and counted the number of larvae, our proxy for mating success, where few or no larvae indicate mating disruption success.
Objective 3. Conduct open-field trials of single-component pheromone blends to disrupt mating and provide crop protection.
Experiment 3. Testing single-component blends to disrupt mating and reduce crop damage in the field.
We tested whether chiral and racemic single-component pheromone treatments (either 100 ug 2S,10S-diacetoxyundecane or 400 ug 2,10-diacetoxyundecane, respectively) successfully confused males and provided protection of a broccoli test crop. We used 80 polyethylene bag pheromone dispensers (Fig. 3) to distribute the pheromone in 52x52 ft plots of broccoli (described for field calling experiments). Plots were situated at least 1,640 ft away from each other to avoid contamination of pheromone treatments via wind. We installed the dispensers only once, and did not replace them during the season. The broccoli was managed organically without any insecticides and grown according to regional practices. Our field plots were located at the University of Guelph Elora Research Station, Pfenning's Organic Farm, a commercial organic vegetable operation in New Hamburg, ON, and at the Institut de recherche et de développement en agroenvironnement (IRDA) in St-Bruno, QC. We chose these locations because of their well-established populations of swede midge, which allowed us sufficient and dependable swede midge pressure to test our treatments. We had one block (one plot of each treatment) at Elora, Pfenning's, and IRDA in 2017, and one block at Elora in 2018.

We collected data throughout the broccoli's growing season on trap shutdown and swede midge damage to the broccoli. In addition to our pheromone dispensers, each plot also contained four commercial monitoring traps for swede midge. Each trap contained the commercial swede midge lure, which releases an attractive dose of the swede midge pheromone at much lower concentrations than the dispensers (Fig. 7). We checked the traps weekly for males. When pheromone mating disruption is successful, there should be few to no males caught in traps (trap shutdown), indicating that the males are confused and cannot locate attractive pheromone sources. At three and six weeks after transplanting we rated 60 random plants per plot for damage to vegetative parts using a three-point damage scale for presence of swollen petioles, stem scaring, leaf curling, and damage to the growing point. At the time of harvest, we rated 60 broccoli crowns per plot for marketability (either yes or no) and scarring and deformation of the head using a five-point scale of increasing severity.
To determine whether pheromone blend was a predictor of the number of males caught in traps in the plots, we used a GLM with GEE model. We used ordinal logistic regression and Kruskal Wallis tests to compare damage ratings and numbers of marketable broccoli heads across pheromone treatments.
Objective 1. Test the ability of pheromone blends to elicit non-competitive mechanisms of disruption.
Experiment 1.1. Male midge mate-seeking behavior following pheromone pre-exposure.
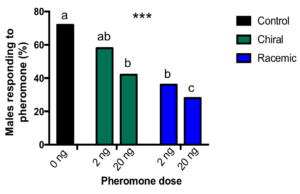
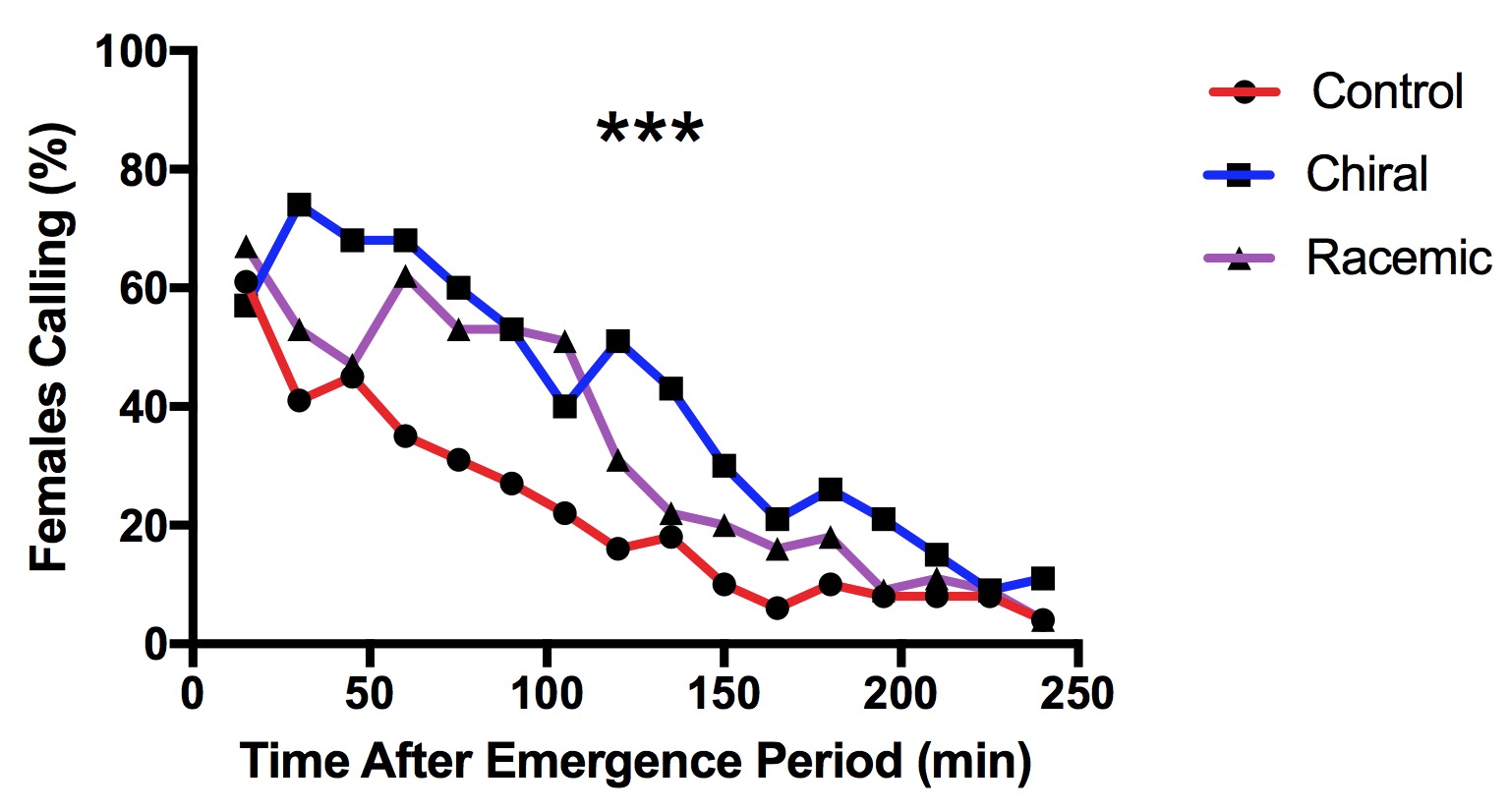
We found that males pre-exposed to both low and high doses of chiral or racemic pheromones were significantly less likely to fly towards female-mimicking attractive pheromones afterward (X32 = 24.882, p<0.001, n=50, Fig. 8). We observed the greatest reduction in mating when midges were exposed to the high dose of racemic pheromone. While 72% of midges located the pheromone when pre-exposed to the solvent-only control, only 28% of midges from the 20 ng racemic treatment flew toward the pheromone. For both chiral and racemic blends, we observed that the effect was dose dependent; fewer males located the pheromones after being exposed to the higher versus lower doses.
Our results indicate that the racemic blend may be more effective in desensitizing male midges, disabling them from locating females in a mating disruption system. However, we also observed a significant reduction in mate-seeking by the chiral blend.
Experiment 1.2. Effect of pheromone pre-exposure on reproductive behavior of females.
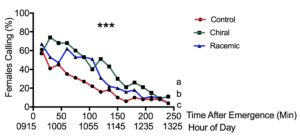
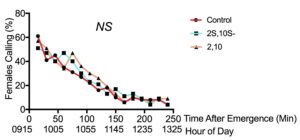
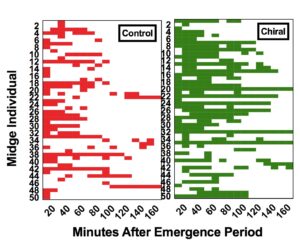
We found that females significantly increased their pheromone-releasing behavior in the laboratory when exposed to synthetic three-component pheromone treatments compared with the solvent-only control (F2,2397=25.586, p<0.001, n=50, Fig. 9). This effect was not observed for the one-component treatments (F2,2397=0.883, p>0.05, n=50, Fig. 10), indicating that the females need at least two out of the three components in their pheromone blend to respond. The effect of increased calling was most pronounced when midges were exposed to the blend they naturally produce, the three-component chiral blend. We observed midges frequently calling for longer periods of time compared with those not exposed to pheromone (Fig. 11).
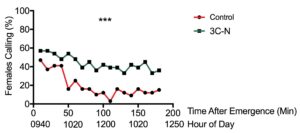
We observed a similar effect in the field; midges were significantly more likely to call in pheromone-treated plots versus untreated plots (X21 = 15.169, p<0.001, n=33, Fig. 12). On average, twice as many midges called in the mating disruption plots at any given time (44.1 ± 1.4% vs. 19.8. ± 2.2% of control midges). Additionally, pheromone exposure significantly influenced the number of continuously-calling midges (X21 = 4.524, p<0.01). Eight times more midges called continuously in the pheromone-treated plot versus (24% of midges vs. 3% in the control treatment).
The ability of females to respond to their own pheromone may be a response that has evolved to help the insects deal with competition from other members of their own species. Females may perceive pheromone in their surroundings as indicators that there are other females around, and may increase their own pheromone releasing behavior to attract mates away from other females. This response on its own may decrease the efficacy of mating disruption if females can attract males by releasing more pheromone. Alternatively, females continuously releasing pheromones may be using limited energy resources, since adult midges do not feed as adults. This study may be the first of its kind to document autodetection in gall midges, and additional research is necessary to elucidate the effects of increased calling on mating disruption efficacy.
In the second part of the experiments, we found that females were significantly less likely to mate following exposure to three-component chiral and racemic pheromones (X22 = 7.354, P <0.05, n=32). While 68% of the midges mated in the control treatment, only 42% and 35% mated in the chiral and racemic treatment groups, respectively. Among the females that did not mate, we did not observe significant differences in calling or female rejection.
Perhaps females were less likely to mate after pheromone pre-exposure because they perceived the pheromones as indicators of many females in the area, and thus competition. While increased calling when exposed to pheromones appears to indicate increased receptivity to mating, decreased propensity to mate following pheromone pre-exposure appears to convey the opposite. It is also possible that pheromones desensitized females to male aphrodisiac pheromones, preventing them from accepting males for mating. However, it is currently unknown whether male midges produce pheromones. Future experiments testing how long pheromone pre-exposure effects last will be useful to determine whether females recover from decreased propensity to mate.
Objective 2. Test the effect of swede midge population density on the efficacy of pheromone blends for mating disruption in enclosures.
Experiment 2. Population density effects on mating disruption efficacy.
We experienced difficulties achieving mating disruption in our small cages. We found that numbers of larvae (our proxy for mating success) between control, chiral, and racemic treatments within each midge density were approximately equal based on n=20 replicates. It is possible that the midges were able to find each other easily because the cages were small, or that the amount of pheromone released from only one dispenser (without coverage from additional dispensers) was not sufficient to confuse males. We were unable to test which pheromone blend was most effective at high midge population densities. However, we feel as though our previous mating disruption experiments in the field (for this grant and our USDA NIFA CPPM grant) allowed us to test which blends were most effective in highly infested areas, due to the large populations of midges at the farms where our plots were located.
Objective 3. Conduct open-field trials of single-component blends to test their ability to disrupt mating and provide crop protection.
Experiment 3. Testing single-component pheromone blends to disrupt mating and reduce crop damage in the field.

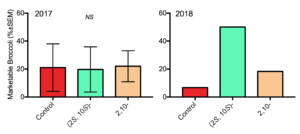
In 2017 and 2018, we obtained data on trap shutdown and crop protection in n=4 field plots treated with the chiral and racemic main pheromone components compared with an untreated control. We found that neither pheromone treatment confused males causing trap shutdown compared with the untreated control (X32 = 1.373, P >0.05, Fig. 13). Little damage was observed at three and six weeks after transplanting in all plots. At the end of the season the numbers of marketable broccoli were not significantly different across treatments (H2 = 0.787, P > 0.05, Fig. 14). Single-component pheromone treatments are not effective for swede midge mating disruption.
 Accomplishments
Accomplishments
Through these projects, we gained insight into the effects of synthetic pheromone mating disruption treatments on male and female midge behavior and the efficacy of lower cost, single-component pheromone treatments for mating disruption. We found that females are capable of detecting and responding to synthetic pheromones, and that they may be less likely to successfully mate following synthetic pheromone pre-exposure. Pre-exposure to three-component chiral and racemic pheromone blends desensitized males and rendered them unable to find female-mimicking pheromones afterwards. We also found that lower cost, simpler pheromone blends containing only the major pheromone component produced by females are not effective for mating disruption. The single-component treatments failed to increase marketable yield compared to the untreated control. These results suggest that three-component pheromone blends (tested under another separate USDA grant) may be the best candidate pheromone blends.
References Cited:
- Miller, J. et al. Mating disruption for the 21st century: Matching technology with mechanism. Environ Entomol 1-27 (2015)
-
Hillbur, Y. et al. Identification of the sex pheromone of the swede midge, Contarinia nasturtii. J Chem Ecol 31, 1807–1828 (2005).
-
Boddum, T. et al. Optimisation of the pheromone blend of the swede midge, Contarinia nasturtii , for monitoring. Pest Manag Sci 65, 851–856 (2009).
- Holdcraft, R. et al. Pheromone autodetection: Evidence and implications. Insects 7, 17 (2016).
Education & outreach activities and participation summary
Participation summary:
We presented results of our swede midge pheromone mating disruption research, as well as other basic information on swede midge, at the Vermont Vegetable and Berry Growers Association Annual Meeting and Northeast Organic Farming Association (NOFA) Vermont Winter Conference in January and February 2017. Here, we presented information to growers on how to identify swede midge and its damage in crop plants, the insect's life cycle, and an update on current recommendations for management and new management strategies in development. We presented preliminary results from our swede midge pheromone mating disruption trials.
To characterize the experience of the NOFA-VT conference attendees with swede midge, their preferences for outreach materials and their willingness to try new management strategies, we conducted a survey of the audience. Of a total of 13 attendee respondents, 11 were from Vermont and 2 were from Québec, and approximately 69% had experience with swede midge on their farms previously. 85% of attendees indicated that factsheets were one of their preferred methods for learning more about insect pest problems.
In winter-spring 2017, we worked with Canadian swede midge expert Dr. Rebecca Hallett, and Cornell University Extension collaborator Christine Hoepting to write a factsheet on organic management of swede midge. This four-page document outlines basic information on swede midge biology, damage symptoms in crops, current recommendations for organic management, and short summaries of strategies currently being researched (including pheromone mating disruption). The factsheet was made available at the NOFA-Vermont Winter Conference in February 2017.
OrganicSMFactsheet_Mar2019PDF
In fall 2017 and 2018, we presented research results on swede midge pheromone mating disruption to academics (including some involved in outreach and extension) at the Entomological Society of Canada (ESA) and Entomological Society of America (ESA) meetings in Winnipeg, MB (ESC), Denver, CO (ESA), and Vancouver, BC (joint ESA/ESC).
In December 2017, shared results of our pheromone mating disruption trials at Les Journées Horticoles et Grandes Cultures conference in St-Rémi, Québec to Canadian vegetable growers and agricultural service providers. At this meeting we were able to network with potential future collaborators and learn about how our Canadian neighbors are managing swede midge. In January 2018, we participated in a videoconference in L'Assomption, QC and gave the same presentation, which was broadcast in 15 locations around Quebec so that growers could attend in their region.
Project Outcomes
We have interacted with many growers in both the U.S. and Canada who are very interested in pheromone mating disruption for swede midge because they have few other options to manage this challenging pest. For example, one grower emphasized that although he is currently spraying his broccoli weekly for swede midge and that it is effective for reducing damage, he is concerned that the insects may be developing resistance. Another grower lamented that they have reduced their Brassica acreage since experiencing swede midge damage because they have no suitable options for control and are losing money. Both were highly interested in mating disruption. Growers in the Northeast are clearly interested in pheromone mating disruption because it limits impacts on non-target organisms and allows them to reduce insecticide use, and may allow them to continue growing Brassica crops effectively. Broccoli, turnips, kale, and other Brassica crops are staples in northern climates because they are cold tolerant. Our results have helped further the development of an organic management practice that may someday allow growers to continue producing these valuable crops profitably.
The most significant outcomes of our research are that we believe we have identified the most promising pheromone blend for swede midge mating disruption and better understand how midges respond to synthetic pheromones. We have identified the next necessary steps in the research and development of a commercial swede midge mating disruption system, which allowed Dr. Yolanda Chen to acquire a SARE Novel Approaches grant (LNE18-368R), titled, "Potential for a pheromone mating disruption program for the invasive swede midge in complex annual rotational systems." This grant will pursue understanding where midges mate in annual cropping systems, as well as further elucidate competitive versus non-competitive mating disruption mechanisms by testing varying pheromone dispenser densities. Knowing where midges mate and whether few or many dispensers are needed to confuse males will inform the location and densities of dispensers in future mating disruption systems.