Final report for GNE21-258
Project Information
Fire blight is a devastating disease of apple and pear caused by the bacterium Erwinia amylovora (Ea). Control of the disease is primarily achieved through antibiotic spraying, which while effective, can drive evolution of antibiotic-resistant Ea strains. A complimentary management strategy involves developing varieties of apple and pear with genetic resistance to fire blight. However, progress in this area has been slow to date, and most varieties of apple and pear remain highly susceptible to fire blight. In this proposal, we seek to identify additional genetic sources of resistance that can potentially be deployed in apple and pear to combat fire blight.
HrpN is a proteinaceous virulence factor secreted by Ea that is essential for disease development. Tobacco, a fire blight nonhost, gives a rapid and powerful defense response to purified HrpN injection into leaves. This suggests tobacco has a yet-undiscovered immunity gene that is recognizing HrpN and triggering a defense response. To uncover the identity of this gene, we took a genetic mapping approach through the crossing of a HrpN unresponsive and a HrpN responsive tobacco accession. Segregation of the defense response trait suggested a single, dominant locus was mediating the response. To identify candidate genes in this locus, we performed whole-genome sequencing on two pooled DNA samples of 2nd generation individuals, one consisting of HrpN-responsive plants and one from HrpN-unresponsive plants. Analysis of the sequencing data suggested the locus is on chromosome 2 in the tobacco genome. We identified several candidate defense genes on chromosome 2, many with receptor-like characteristics.
Ongoing work in our lab will test these candidates for their involvement in HrpN perception, particularly if any of these candidates function as a HrpN receptor. Apple does not respond to purified HrpN, so we are excited by the prospect of transferring one or more gene candidates to apple, where they could potentially sense Ea-delivered HrpN and institute a defense response. Overall, this project was successful in elucidating the genetics of HrpN perception in tobacco. Continued research has the potential to contribute novel sources of fire blight resistance to the agricultural community.
Objective 1: Determine the inheritance patterns of the HrpN-induced HR in one or more mapping populations of tobacco.
Objective 2: Identify one or more candidate resistance genes in a tobacco mapping population by performing a bulked segregant analysis.
Fire blight, caused by the bacterium Erwinia amylovora (Ea), is a devastating disease that primarily afflicts apple (Malus x domestica) and pear (Pyrus communis). The disease is characterized by wilting and death of flowers and shoots, production of bacterial ooze from plant parts, and canker formation on the tree trunk. Left unmanaged, fire blight can result in death of the entire tree. Outbreaks of fire blight in orchards are costly to growers, often resulting in millions of dollars in crop and tree loses (van der Zwet, Orolaza-Halbrendt, and Zeller 2012). Treatment of fire blight currently relies on application of antibiotics (e.g. streptomycin). However, heavy antibiotic use has led to the rise of streptomycin-resistant Ea strains that are causing recent outbreaks in the Northeastern US (Dougherty et al. 2021; Russo et al. 2008). Additionally, antibiotic application to flowers must be correctly timed, as antibiotics are not effective once Ea has gained access to internal plant tissues. This suggests the need to develop genetic sources of resistance as a complementary fire blight management strategy. Currently no apple cultivars with complete resistance to fire blight are available, and many of the most economically important apple and pear cultivars are highly susceptible to fire blight (van der Zwet, Orolaza-Halbrendt, and Zeller 2012; Kostick, Norelli, and Evans 2019).
HrpN is a proteinaceous virulence factor secreted by Ea that facilitates the bacterium’s infection of plant tissues (Wei et al. 1992). While HrpN functions in fire blight development on host plants, it has been shown to activate plant defenses in nonhost plants. HrpN, when secreted by Ea or infiltrated as a cell-free elicitor, induces a strong, rapid, and localized cell death defense reaction called the hypersensitive response (HR) in leaves of the nonhost plant Nicotiana tabacum (cultivated tobacco). The HR is associated with gene-for-gene recognition and plant disease resistance, leading us to hypothesize that tobacco possesses a HrpN-recognizing resistance (R) protein.
Through the objectives proposed in this study, we have identified several tobacco candidate genes that facilitate HrpN immune recognition. Interestingly, apple does not mount an immune response to purified HrpN (Reboutier et al., 2007). Upon further testing, one or more of our candidate genes could be transferred to apple and pear to potentially confer HrpN recognition and increase fire blight resistance. The long-term goal of this project, development of fire blight-resistant apple and pear, will directly assist Northeast farmers by increasing productivity, reducing costs, and increasing net farm income. Crop losses due to fire blight outbreaks will decrease, increasing the productivity and profitability of grower operations. Additionally, growers will save on costs associated with annual chemical treatments, including product, labor, and transportation costs. These reduced costs will ensure that healthful produce is available to consumers at a reasonable price. Additionally, lowered antibiotic use provides a reduction of environmental risks in agriculture by reducing the prevalence of antibiotic-resistant microbes and avoiding off-target effects on beneficial microbes.
References
Dougherty, Laura, Anna Wallis, Kerik Cox, Gan-Yuan Zhong, and Benjamin Gutierrez. 2021. “Phenotypic Evaluation of Fire Blight Outbreak in the USDA Malus Collection.” Agronomy 11 (1): 144
Kostick, S. A., J. L. Norelli, and K. M. Evans. 2019. “Novel Metrics to Classify Fire Blight Resistance of 94 Apple Cultivars.” Plant Pathology 68 (5): 985–96.
Reboutier, David, Cécile Frankart, Joël Briand, Bernadette Biligui, Sandrine Laroche, Jean-Pierre Rona, Marie-Anne Barny, and François Bouteau. 2007. “The HrpNEa Harpin from Erwinia amylovora Triggers Differential Responses on the Nonhost Arabidopsis thaliana Cells and on the Host Apple Cells.” Molecular Plant-Microbe Interactions 20 (1): 94–100.
Russo, Nicole L., Thomas J. Burr, Deborah I. Breth, and Herb S. Aldwinckle. 2008. “Isolation of Streptomycin-Resistant Isolates of Erwinia amylovora in New York.” Plant Disease 92 (5): 714–18.
van der Zwet, Tom, Noemi Orolaza-Halbrendt, and Wolfgang Zeller. 2012. Fire Blight: History, Biology, and Management. St. Paul, USA: American Phytopathological Society.
Wei, Zhong-Min, Ron J. Laby, Cathy H. Zumoff, David W. Bauer, Sheng Yang He, Alan Collmer, and Steven V. Beer. 1992. “Harpin, Elicitor of the Hypersensitive Response Produced by the Plant Pathogen Erwinia amylovora.” Science 257 (5066): 85–88.
Research
Objective 1: Determine the inheritance patterns of the HrpN-induced HR in one or more mapping populations of tobacco.
Rationale
The strong cell death defense response induced by HrpN in tobacco is suggestive of R gene mediated recognition. As tobacco is a diverse crop species cultivated throughout the world, I can screen these accessions for natural variation in the HrpN response. A small number of accessions have likely accumulated genetic mutations in the HrpN-recognizing R gene through random human-mediated selection processes. Those accessions with a defective receptor will show a weak or no cell death response when its leaves are infiltrated with HrpN protein.
In preliminary studies, I screened 127 tobacco accessions collected from around the world by infiltrating their leaves with HrpN protein and visually assessing the cell death response. I identified 12 tobacco accessions that were deficient in their visual cell death response to HrpN. I then used an electrolyte leakage assay to identify three accessions with the quantitatively weakest cell death response to HrpN. The three unresponsive tobacco accessions likely have genetic mutations in one or more genes that are responsible for the HrpN-induced HR. Crossing of these accessions with a HrpN-responsive accession will help elucidate the heritability and segregation patterns of the trait.
Methods
F1 Generation and Phenotyping
To generate F1 populations, each unresponsive tobacco accession will be crossed with the HrpN-responsive tobacco accession TI 1608. To perform a cross, anthers are removed from an unopened flower that is within 24 hours of opening. The emasculated flower’s stigma is then immediately pollinated with an anther from the proper accession. 4 weeks after pollination, the seed pods are removed, and the seed is collected and stored in manila envelopes.
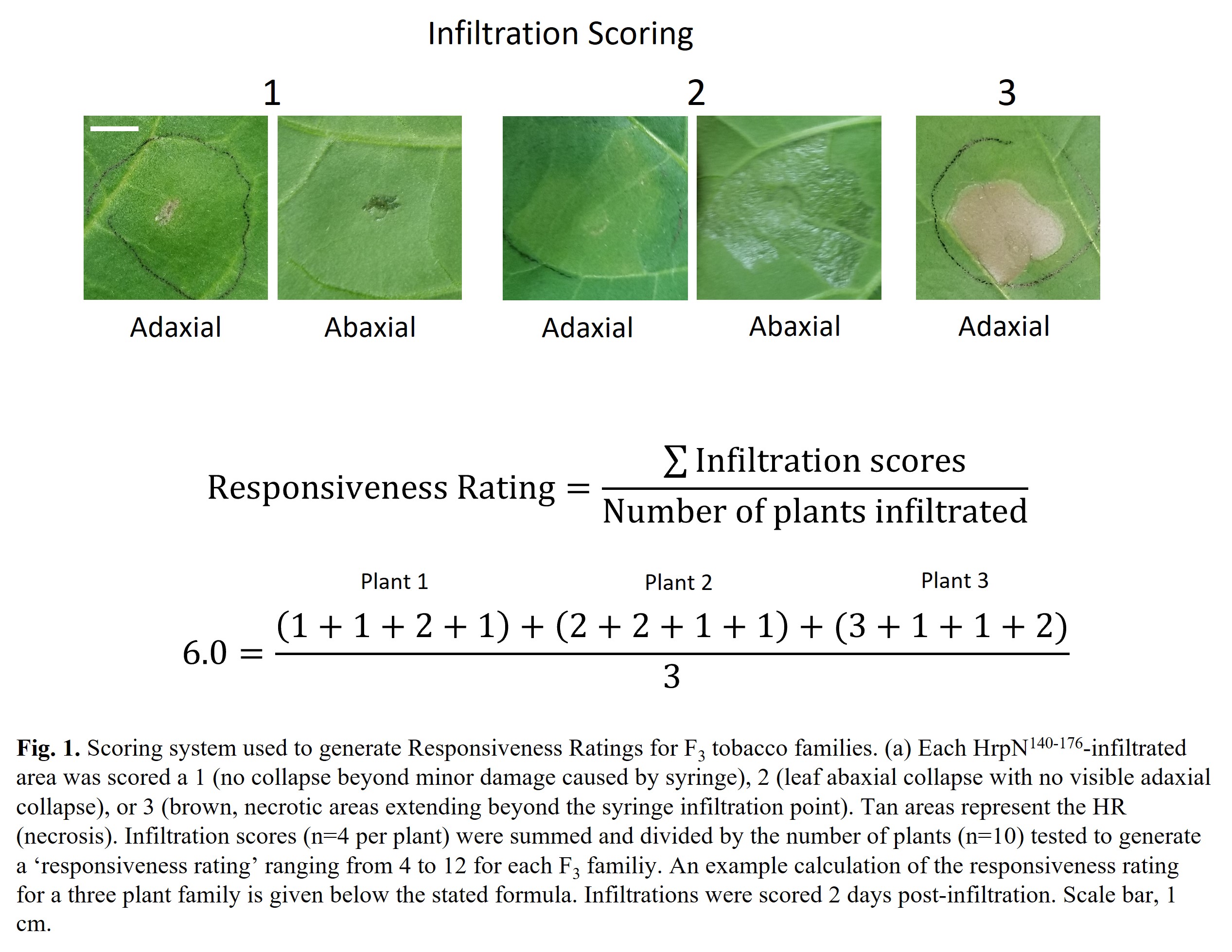
Once seed has been collected, 40 individuals of each of the three F1 families will be grown to 6 weeks old in a plant growth chamber. Four leaves on each plant will be infiltrated with 100 µM HrpN142-188, a peptide corresponding to amino acids 142 to 188 of full-length HrpN protein. This peptide represents the major HR-inducing segment of HrpN. An advantage of using HrpN142-188 is that it is the simplest HR trigger available to test HrpN-mediated plant responses and eliminates other potential plant reactions triggered by full-length HrpN. Two days after infiltration, leaves are scored visually using a rating scale: 1 (no tissue necrosis), 2 (mild underside tissue collapse), or 3 (complete tissue necrosis). Each individual is then classified as strongly responsive (≥ 75% of infiltrations scored a 3), unresponsive (≥ 50% of infiltrations scored a 1), or moderately responsive (not meeting the criteria of strong or weak responders). As R gene mediated recognition of pathogen virulence factors is typically a dominant trait, it is expected that all individuals in each population will be strongly or moderately responsive to HrpN142-188.
01/22 Updates
The HrpN-responsive tobacco accession used in F1 generation was changed to TC 319. This accession has a chromosome-level genome assembly (Sierro et al. 2024). The use of TC 319 will lead to a stronger sequencing data analysis in Objective 2. TC 319 has been verified to respond to HrpN peptide.
The peptide HrpN142-188 has been replaced with HrpN140-176. This peptide has increased potency from HrpN142-188 and represents a more specific trigger of HR in tobacco.
For scoring of progeny in the F1 and F2, a binary classification system was adopted. Individuals were classified as responsive if at least one infiltration resulted in a score of 3 (complete tissue necrosis). Individuals with infiltrations resulting in only scores of 1 and 2 are classified as unresponsive. This will allow for easier grouping of individuals into HrpN-responsive and HrpN-unresponsive pools as outlined in Objective 2.
F2 Generation and Phenotyping
Plants in each of the F1 populations will be allowed to self-pollinate. Seed will be collected from the selfed plants as described previously. 200 individuals of each of F2 population will be grown and phenotyped with HrpN142-188 using the same methods as the F1 screen. To accommodate the large number of plants, some plants may be grown in rented space in a university greenhouse. In the F2 populations, a segregation of phenotypes is expected, where some plants are strongly or moderately responsive and others are unresponsive. A chi-squared test will be used to determine the segregation patterns of each population. The F2 which most closely resembles a Mendelian segregation pattern (e.g. 3:1) of strongly/moderately responsive: unresponsive individuals will be utilized for the bulked segregant analysis (BSA), as this indicates the trait is controlled by a small number of genes. However, the BSA may also be utilized in populations displaying more complex inheritance patterns, as long as individuals are sorted into contrasting phenotypic pools (see Objective 2).
01/22 Updates
To confirm the HrpN-responsive and HrpN-unresponsive designations of F2 individuals, each plant will be selfed to generate F3 populations. 10 individuals from each F3 population will be screened with HrpN140-176 and classified as HrpN responsive or unresponsive. F3 populations that contain no trait segregation (i.e. consisting of individuals only HrpN responsive or only HrpN unresponsive) will be utilized in the BSA, while F3 populations displaying trait segregation (indicating a heterozygous F2 parent) will be excluded. This will increase the confidence in assigning F2 individuals to HrpN responsive and unresponsive groups and lead to more effective analysis of the whole-genome sequencing data (see Objective 2).
01/24 Updates
To better evaluate the responsiveness of the F3 families to HrpN140-176, a 'Responsiveness Rating' was calculated for each family. Each individual was leaf infiltrated four times with HrpN140-176. After two days, each infiltration was scored using the 1,2,3 system described above. These values were then summed for an individual, giving a responsiveness rating between 4 and 12. Finally, the individual ratings were averaged across the 10 screened F3 individuals to give an overall responsiveness rating for each F3 family. See Fig. 1 below for example calculation of responsiveness ratings.
Objective 2: Identify one or more candidate resistance genes in a tobacco mapping population by performing a bulked segregant analysis.
Rationale
Once the segregation patterns have been determined in my F2 population, a bulked segregant analysis (BSA) can be performed. A BSA consists of pooled DNA samples from a segregating population (e.g. my F2 population), where each pool corresponds to a specific phenotype. These pools are then subjected to whole-genome sequencing. The pooling of many individuals in each sample allows the investigator to identify genetic mutations, typically single nucleotide polymorphisms (SNPs), that are strongly associated with the phenotype of interest. Additionally, the pooling of samples, rather than sequencing of individuals, saves sequencing costs and simplifies analysis.
Methods
Sample Grouping and DNA Extraction
F2 individuals will be grouped (i.e. bulked) into two phenotypic categories: HrpN-responsive (consisting of individuals identified as strongly or moderately responsive in Objective 1) or HrpN-unresponsive. DNA will be extracted individually from leaf tissue of 30-40 individuals in each phenotypic group using the Qiagen DNeasy Plant Mini kit. DNA quality will be assayed using a department-provided NanoDrop. For each group, the combined DNA sample will be created by mixing 2 µl of each individual sample.
DNA Sequencing
The two pooled DNA samples will be submitted to the Penn State Genomics Core Facility for library construction and DNA sequencing. Libraries will be assembled using the Illumina DNA PCR-free prep kit. Genome sequencing will be done using two runs of NextSeq High Output 150x150 paired-end sequencing. As the tobacco genome is 4.5 gigabases, this will generate ~30x coverage of reads across the genome. 30x coverage provides sufficient sequencing data to detect genetic variation between the phenotypic groups (Schultink et al. 2019).
Data Analysis
Computational analyses will be conducted using the Penn State’s Roar supercomputer, a free high-performance research cloud. Reads will be mapped to the tobacco reference genome (Sierro et al. 2024) using BWA-MEM (Li and Durbin 2010). Calling of SNPs and other genetic polymorphisms will be performed using GATK (McKenna et al. 2010). BWA-MEM and GATK are freely available software that are commonly employed in the analysis of DNA sequencing data. SNPs and other polymorphisms will be filtered based on the following criteria: a differential abundance of at least 25% between the two pools, a minimum mapping quality of 10, and their presence in the coding sequence or promoter of genes with structural features of known defense genes (e.g. receptor-like kinases and receptor-like proteins). The genome location of polymorphisms which meet these criteria will be used to identify up to 10 high quality gene candidates that may be mediating tobacco’s defense response to HrpN.
A graphical overview of Objectives 1 & 2 can be found here: Experiment Diagram
References
Li, Heng, and Richard Durbin. 2010. “Fast and Accurate Long-Read Alignment with Burrows-Wheeler Transform.” Bioinformatics 26 (5): 589–95.
McKenna, A., M. Hanna, E. Banks, A. Sivachenko, K. Cibulskis, A. Kernytsky, K. Garimella, et al. 2010. “The Genome Analysis Toolkit: A MapReduce Framework for Analyzing next-Generation DNA Sequencing Data.” Genome Research 20 (9): 1297–1303.
Schultink, Alex, Tiancong Qi, Julia Bally, and Brian Staskawicz. 2019. “Using Forward Genetics in Nicotiana benthamiana to Uncover the Immune Signaling Pathway Mediating Recognition of the Xanthomonas perforans Effector XopJ4.” New Phytologist 221 (2): 1001–9.
Sierro N, Auberson M, Dulize R, Ivanov N V. 2024. Chromosome-level genome assemblies of Nicotiana tabacum, Nicotiana sylvestris, and Nicotiana tomentosiformis. Scientific Data 11: 135.
Objective 1
F1 generation and phenotyping
Four F1 families, representing crosses of different HrpN-unresponsive accessions with the responsive accession TC 319, were screened with HrpN140-176. In the TC 319 x TI 126 cross, every individual tested was rated as HrpN responsive (Table 1), suggesting that the HrpN response has a genetic basis and is a dominant trait.

F2 Generation and Phenotyping
An F1 individual from the TC 319 x TI 126 cross was selfed to produce an F2 population. 220 F2 individuals have been screened with HrpN140-176. Of these, 174 individuals were classified as HrpN responsive and 46 as HrpN unresponsive. This result is in line with an expected 3:1 ratio of HrpN responsive:HrpN unresponsive individuals (Chi-sqaure probability=0.16). Thus, HrpN recognition appears to be governed by a single gene.
Approximately 160 F2 individuals are currently being grown in a greenhouse for seed collection. Screening of the F3 lines from these individuals is expected to conclude in summer 2022.
01/23 Update-Objective 1
A total of 346 F2 individuals have now been screened with HrpN140-176. These results are supportive of a 3:1 ratio of HrpN responsive:HrpN unresponsive individuals (Chi-square probability=0.85), suggesting a single, dominant locus (Table 2).

I have screened 64 F3 families-31 from responsive F2 parents and 33 from unresponsive F2 parents. Of these, 18 and 16 F3 families had consistent phenotypes with their responsive and unresponsive F2 parent phenotypes, respectively. Thus, these F2 parents will be utilized in the bulked segregant analysis. I am currently in the process of screening 29 more F3 families from responsive F2 parents and 27 more from unresponsive F2 parents. At the current rate, I will have at least 25 responsive F2 parents and 25 unresponsive F2 parents that can be utilized for sequencing by early Spring. One more round of F2/F3 screening may be needed to ensure adequate numbers of individuals in each phenotypic category.
01/24 Update-Objective 1
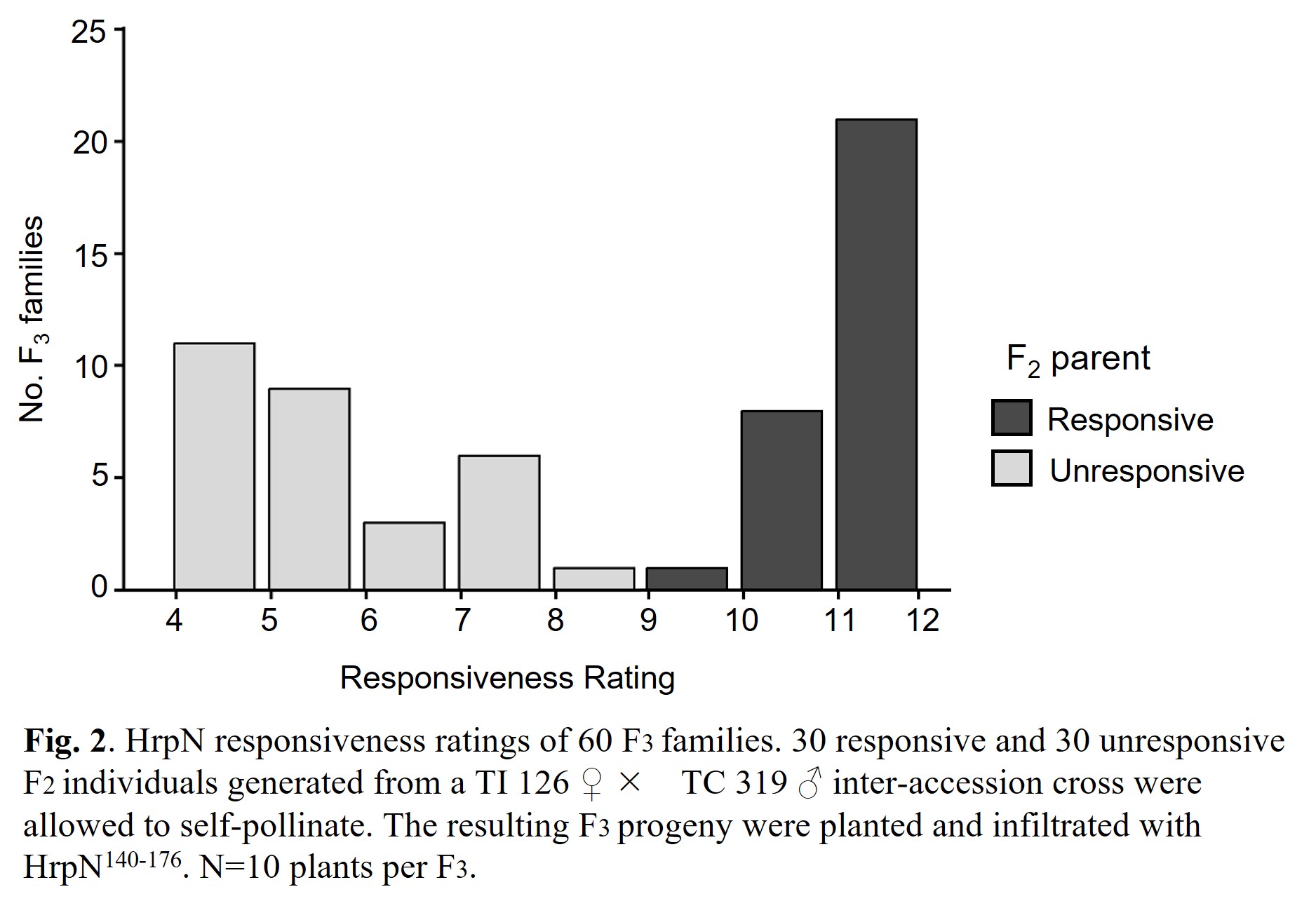
F3 screening concluded in spring 2023. A total of 113 F3 families were screened- 53 from responsive F2 parents and 60 from unresponsive F2 parents. For each F3 family, a 'responsiveness rating' to HrpN140-176 was calculated (Fig. 1).
01/24 Update-Objective 2
Sample Grouping
The results of the F3 phenotyping were used to inform selection of F2 parents to be used in sequencing. For sequencing, a minimum of 30 F2 individuals of each phenotypic class (HrpN-responsive and HrpN-unresponsive) were needed. Using the F3 responsiveness ratings, we identified 30 F2 individuals in each category with progeny that gave a consistent phenotype. F3 families from the chosen responsive F2 parents had an average responsiveness rating of 11.4, while F3 families from the chosen unresponsive F2 parents had an average responsiveness rating of 5.6 (Fig. 2). Thus, we have increased confidence in the phenotypic assignments of our F2 individuals because the trait showed strong heritability into the next generation.
DNA Extraction
Young leaf tissue (stored at -80 C) from the 60 F2 individuals chosen for sequencing was utilized for DNA extraction. All extractions were performed with the Promega Wizard Genomic DNA Purification Kit. The amount of DNA in each extraction was determined through NanoDrop and Qubit assays. Approximately 10 ng of DNA from each of the 30 individual preps in each phenotypic class were pooled to create the responsive and unresponsive DNA samples.
DNA Sequencing
Whole-genome sequencing of the two samples was performed in Summer 2023 at the Huck Genomics Core Facility at Penn State. 150 base pair Illumina paired-end sequencing was utilized. Based on the 4.5 billion base pair size of the tobacco genome (Sierro et al. 2024), we achieved ~40X coverage for each sample (Table 3). This is well above our target of 30X coverage.

Data Analysis
The reads of each sample were mapped to the tobacco reference genome assemble (Sierro et al. 2024). Each sample had ~99% of reads align to the genome assembly. Next, short DNA polymorphisms like single-nucleotide polymorphisms (SNPs), insertions, and deletions were called in each sample, and the effect of each polymorphism (e.g. missense mutation) was annotated. In total, ~11 million DNA polymorphisms were identified across the two samples. Only ~112,000 (1 %) of these are protein-coding altering variants, while the vast majority of the variants are located in intergenic space. The protein-coding altering variants are of interest due to their potential to alter plant traits.
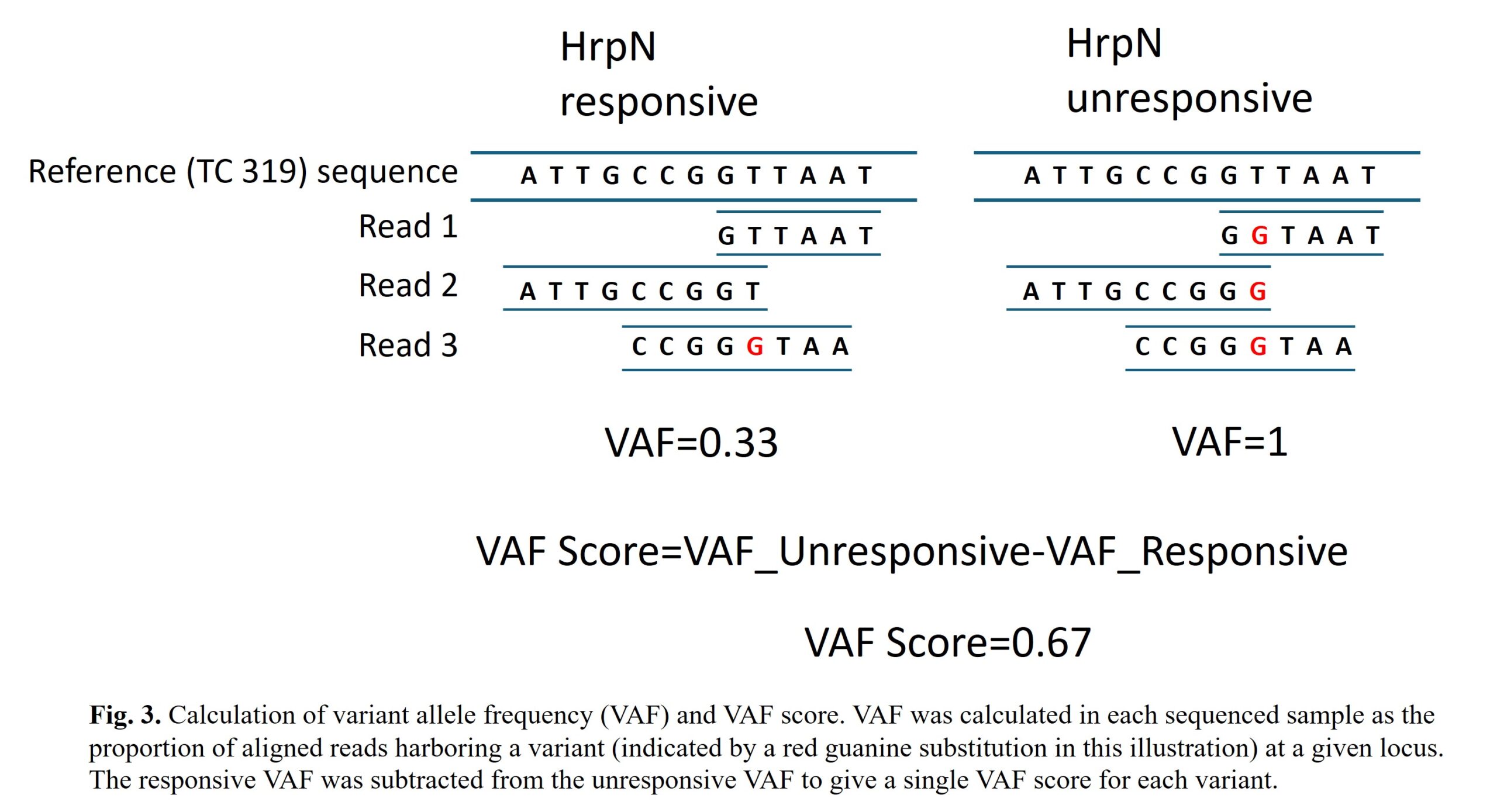
To identify variants that could underlie our trait of interest, we calculated a variant allele frequency (VAF) for all DNA variants in each sample, based on the percentage of sequencing reads that are supporting a given variant (versus ones that support the reference sequence). The VAF calculated in each sample is then subtracted to get a single metric, VAF Score, in which positive values indicate variants associated with unresponsiveness (see example calculation in Fig. 3 below). Because the reference genome of tobacco is derived from the same variety (TC 319) used as the responsive parent in our mapping population, we expect to find high-abundance variants in the unresponsive sample that may alter the function of the HrpN response locus.
Identifying candidate loci
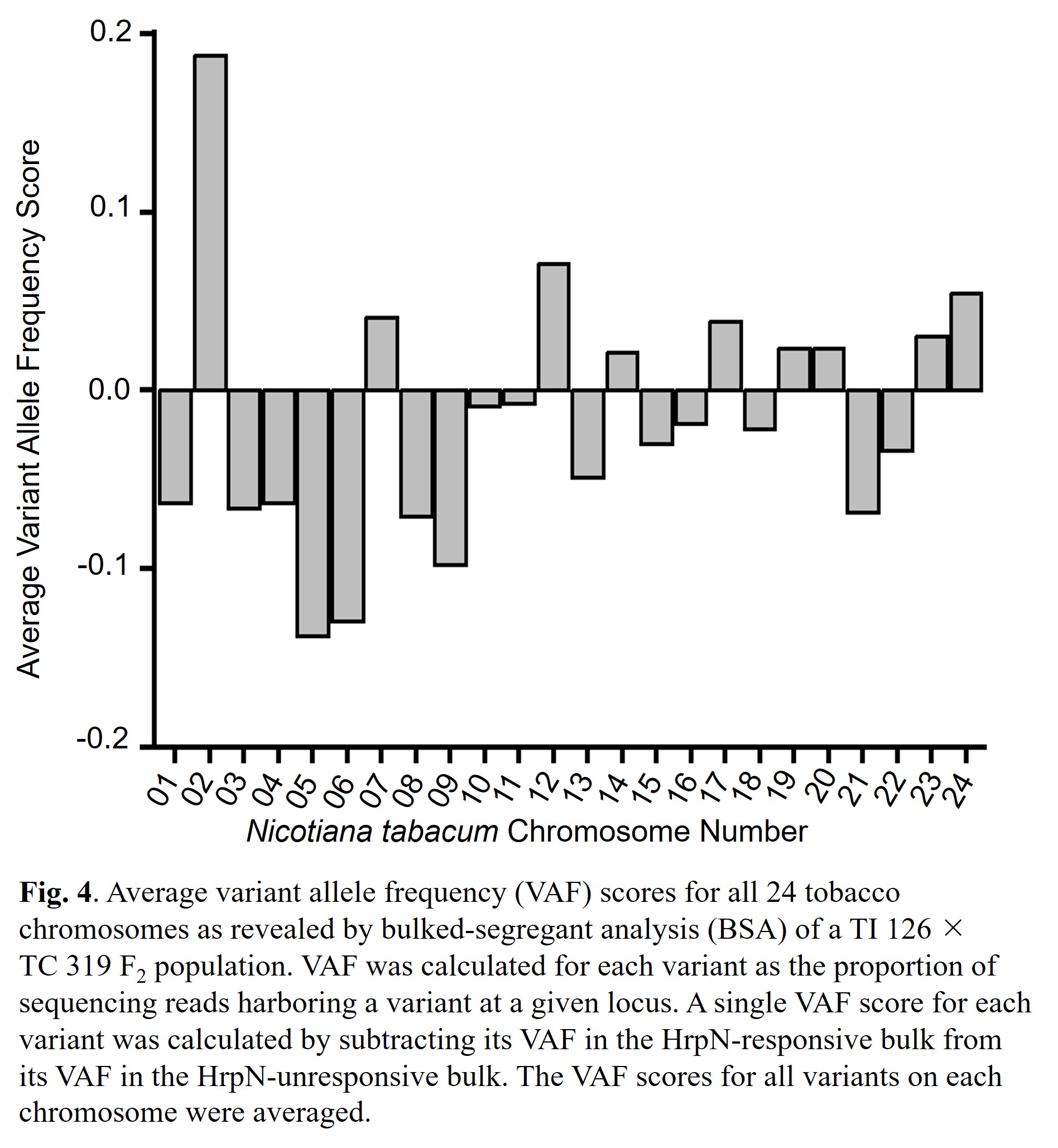
We calculated a VAF score for all identified sequence variants and calculated an average VAF score for all 24 of tobacco's chromosomes. We detected a high VAF score on chromosome 2, indicating our mapped HrpN response locus may lie on chromosome 2 (Fig. 4).
Next, we plotted the locations of protein-coding altering variants along chromosome 2 (Fig. 5). Because we are looking for a defense-related gene, we identified candidate genes annotated as protein kinases. We selected a total of 9 candidate genes, 3 of which are predicted to encode signaling kinases and 6 of which are predicted to encode receptor-like kinases (Fig. 5; Table 4).
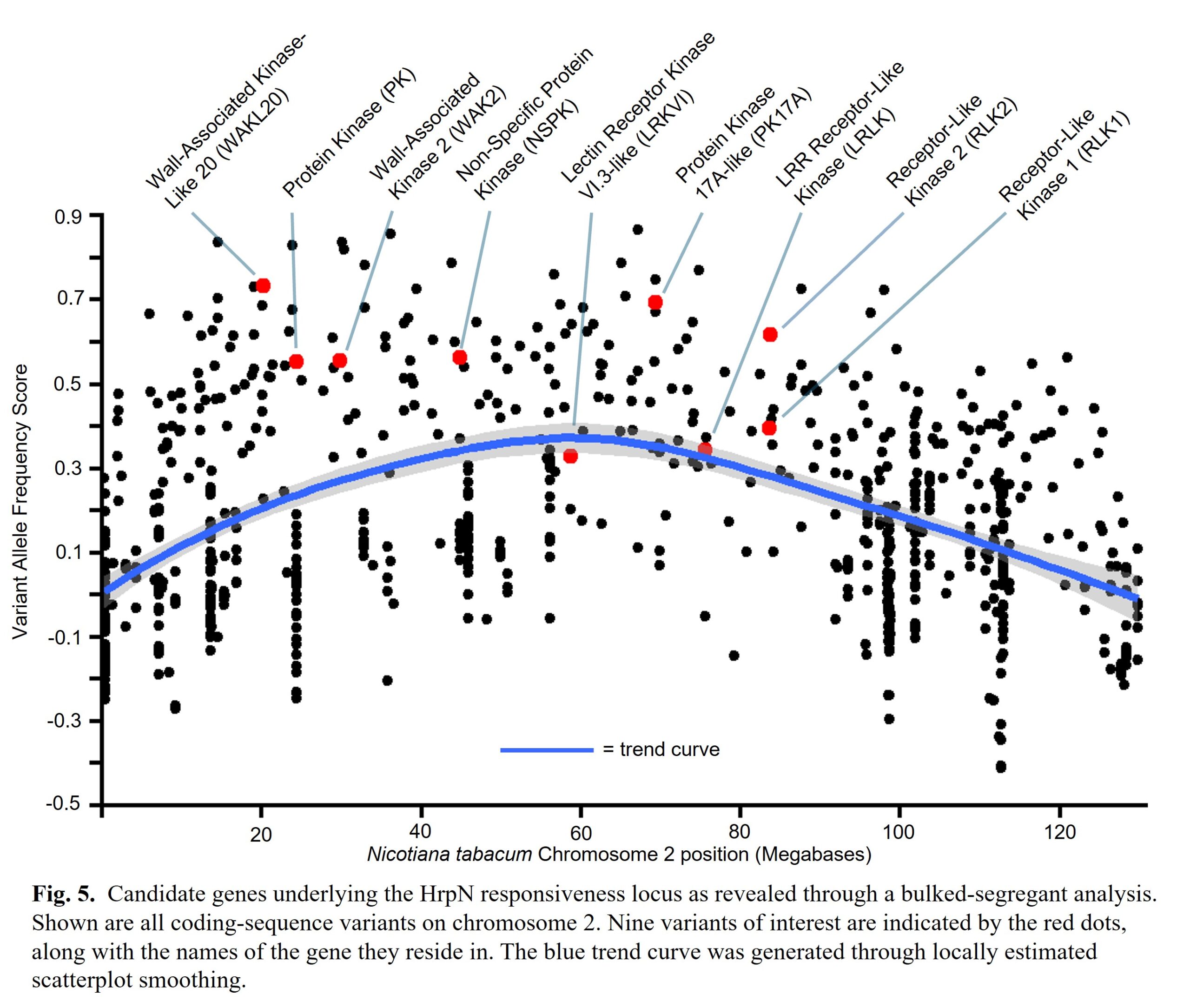

Receptor-like kinases (RLKs) are common immunity proteins that recognize other pathogen molecules, such as flagellin and chitin (Dodds & Rathjen, 2010) . Thus, we are excited by the possibility of one of these candidates functioning in HrpN perception. Our lab has also recently identified wall-asscociated kinase 3 (WAK3) as being genetically required for HrpN perception in the model plant Arabidopsis thaliana (Held et al., 2024). The identification of several WAKs in this dataset could suggest a WAK is also involved in HrpN perception in tobacco. Further testing in our lab will clarify the role of these candidates in HrpN immune recognition in tobacco.
References
Dodds PN, Rathjen JP. 2010. Plant immunity: towards an integrated view of plant-pathogen interactions. Nature Reviews Genetics 11: 539–548.
Held JB, Rowles T, Schulz W, McNellis TW. 2024. Arabidopsis Wall-Associated Kinase 3 is required for harpin-activated immune responses. New Phytologist 242: 853–857.
Sierro N, Auberson M, Dulize R, Ivanov N V. 2024. Chromosome-level genome assemblies of Nicotiana tabacum, Nicotiana sylvestris, and Nicotiana tomentosiformis. Scientific Data 11: 135.
This research was very successful, as we met both proposed objectives. In objective 1, we generated a tobacco mapping population through the crossing of a HrpN-responsive and HrpN-unresponsive accession. The segregation patterns suggested a single, dominant locus was mediating the trait. In objective 2, we conducted whole genome sequencing on pooled F2 individuals, and used bioinformatic analyses to identify nine quality candidate genes. Many of these genes are predicted to encode receptor-like kinases, a major class of immunity proteins.
One major conclusion of this research is that HrpN responsiveness is a genetically-tractable trait in tobacco. This is a historical finding as HrpN's strong immune response in tobacco was discovered over 30 years ago, but its genetic underpinnings have remained a mystery. If we are able to confirm the function of one or more of our candidates (e.g. as a HrpN receptor), this could lead to applications in fire blight disease resistance.
The success of this proposal suggests that nonhosts can be important reservoirs of immunity in fighting disease. This approach of dissecting nonhost immune recognition to pathogen molecules can be applied to diverse pathogen systems.
Education & outreach activities and participation summary
Participation summary:
Results from this work were disseminated locally and nationally. This past year, I presented seminars to my graduate program (Plant Biology) and department (Plant Pathology & Environmental Microbiology) here at Penn State. In August 2022, I presented a poster on this research at the national APS meeting (Plant Health) in Pittsburgh, PA. The poster is attached below. This audience included local researchers in academia, government, extension, and industry. Sharing of my research here helped illustrate the potential of utilizing nonhost species for resistance genes to other plant disease researchers.
We are currently writing up the results from this study for publication in Molecular Plant Pathology. This is a high-quality and fully open-access molecular plant pathology journal. This will reach other plant pathology researchers across the world. Once the paper is accepted, we plan on generating a popular press/news release article, where we highlight both the historical aspects of the research (HrpN perception in plants has been a 30 year old mystery) and its potential practical impacts for fire blight management. An appropriate venue for this article could be APS's Phytopathology News or Penn State's extension newsletter.
The opportunities for outreach will expand past the duration of this project. I will be giving a seminar at Cornell University in April of this year. My work will also be presented at the fourth International Symposium of Fire Blight on Rosaceous Plants to be held in summer 2025. This is the premier conference for fire blight-related research, drawing in hundreds of researchers from around the world. My work will be of value to government and extension personnel who are interested in new ways to combat fire blight. This conference could help open the door to collaborations in future efforts to generate fire blight-resistant apple and pear germplasm.
The long term outreach opportunities for this research are immense. New apple and pear varieties could be generated containing a tobacco resistance gene. Work with other researchers, extension personnel, and enthusiastic growers will be necessary to conduct field trials of the new varieties to test for fire blight resistance and productivity. If trials are successful, these varieties could be marketed to northeast growers as fire blight-resistant germplasm. Additionally, the results of new research spurred by these results will continue to be disseminated through conference presentations and manuscript publication.
Project Outcomes
Edited to Add (June 2025) Additional Publication: https://bsppjournals.onlinelibrary.wiley.com/doi/10.1111/mpp.70096
--------
As apple and pear are clonally-propogated species, breeding them for disease resistance is extremely difficult as the agronomic characteristics (i.e. fruit quality) of the particular variety are lost in the breeding process. Thus, genetic resistance must be achieved primarily through plant transformation techniques. In particular, immunity genes that recognize components of pathogens are high-value targets for transformation. This research has identified candidate genes that may confer recognition to HrpN, a major virulence factor of the fire blight pathogen Erwinia amylovora (Ea). Apple or pear trees transformed with one or more of these genes may be able to recognize HrpN when it is delivered by Ea. This would trigger a defense response in those invaded plant cells, which could potentially limit further Ea spread and reduce the prevalence and/or intensity of fire blight in orchards. The development of fire blight-resistant apple and pear would undoubtedly benefit farmers economically by reducing plant and crop losses due to fire blight outbreaks.
Additionally, the use of genetic resistance mechanisms would potentially mean less annual reliance on chemical treatments (e.g. antibiotics). Currently, streptomycin spraying at bloom time is the major control mechanism of fire blight, and is a substantial annual cost for growers. This has also led to the occurrence of streptomycin-resistant Ea strains. Finally, antibiotic spraying has off-target effects on beneficial microbes. Thus, reduced antibiotic spraying would yield economic and environmental benefits for farmers.
Any research involving plant breeding and phenotyping is time consuming and difficult. We expected our mapping to take ~1 year, but it actually took over 2.5 years. Even with a relatively simple phenotype, cell death, and the use of an herbaceous species (tobacco) we had to screen large numbers of plants to achieve a result we were confident with. And the effort appears to have paid off, based on our successful identification of gene candidates. But we certainly were humbled in the attention to detail and organization required for this effort.
We are actively pursuing functional studies into our identified gene candidates to see which underlie our HrpN response trait. A revised National Science Foundation proposal is being prepared based on the results of this study. We are excited by the possibility that several of these candidates are predicted to encode receptors, which could provide a mechanistic explanation for HrpN perception. Our lab is developing several methods to test for an interaction between candidates and HrpN protein. We have also begun working with a new collaborator to look at any potential defense function of these candidates by using a bacterial pathogen of tobacco. These results would have implications in the use of candidates in conferring fire blight resistance in apple and pear.