Final report for GNE22-303
Project Information
Halyomorpha halys, or the brown marmorated stink bug, is an invasive insect pest that damages tree fruit and other crops. In the Mid-Atlantic, orchard farmers have reported spraying insecticide up to four times as often to control H. halys during outbreaks, which has severe economic and environmental consequences. Increased biological control, or top-down population pressure on pests by natural enemies, reduces the need for insecticide. Often, the degree to which natural enemies cause mortality to their hosts is dependent on both farm-level habitat variables, such as crop diversity, and broader landscape-level variables, such as percent forest in a 3km radius. This project examines egg parasitism, predation, and stink bug abundances across multiple habitats and spatial scales while also assessing long‑term population trends and grower perceptions of H. halys nearly three decades after it’s initial arrival in New Jersey.
In this project, we deployed and monitored H. halys egg masses and Podisus maculiventris, a beneficial predator stink bug which shares a natural enemy complex, during two field seasons in 2022 and 2023. We deployed colony-laid egg masses on the underside of host plants, retrieved them after 3 days in the field, and reared out stink bug nymphs and egg parasitoids in closed petri dishes. In 2022, we deployed 55 egg masses across 6 sites, and in 2023, we deployed 507 egg masses across 15 sites. We classified eggs into different fates by dissecting them under the microscope and using molecular methods. Egg fates included stink bug hatch, reproductive parasitism (parasitoid hatch), nonreproductive parasitism (unemerged eggs positive for parasitoid DNA), chewing predation, sucking predation, and unknown (unemerged eggs not positive for parasitoid DNA). Next, we ran statistical analyses with the 2022 data to investigate how egg mortality differs in peach, soybean, and forested habitats and statistical analyses with the 2023 data to investigate how land-use around peach orchards predicts egg mortality. The 2023 models included measures of land-use diversity and variables that maximized variance in land-use data among farms.
Results from the 2022 study indicate that habitat plays a major role in egg mortality of stink bugs. Podisus maculiventris had higher parasitism and predation than H. halys in forest and soybean, but not in peach. Halyomorpha halys had higher rates of parasitism and predation when in soybean than when in peaches. However, Trissolcus japonicus—H. halys’ closely co-evolved egg parasitoid from Asia that was first detected in NJ in 2017—only parasitized H. halys eggs in peach and forest habitats. To monitor T. japonicus populations and focus the study on a commodity with historically severe H. halys pest problems, the 2023 study was conducted only in peaches. In this study, H. halys and P. maculiventris faced similar egg mortality, but H. halys experienced greater overall parasitism, and P. maculiventris experienced greater predation. At a landscape scale, H. halys egg mortality was positively correlated with increased suburban habitats. Podisus maculiventris egg mortality was correlated with grassland, soybean, and corn along with forest habitats. At a small, 50m scale, egg mortality of both species was greater when peach blocks bordered mixed blocks of flowering vegetables and other annual crops compared to mowed grass, forest, and other tree fruit habitats. It follows that growers should be encouraged to plant mixed annual, diversified crop blocks adjacent to tree fruit orchards to increase biological control of H. halys.
Trissolcus japonicus was detected on all cooperating peach farms. It parasitized H. halys more than P. maculiventris, contributing meaningfully to biological control while minimally impacting the native beneficial stink bug. This suggests that future T. japonicus releases may be feasible and beneficial for growers to control H. halys populations. Additionally, the widespread growth of T. japonicus populations may be contributing to negative H. halys population trends over time and reduced perceived pest priority as documented by the Rutgers Fruit IPM Program.
Overall, this project demonstrates that biological control of H. halys is shaped by both habitat-scale and landscape-scale factors. The findings can contribute to the advancement of ecologically informed integrated pest management strategies such as habitat management.
The ultimate objective of this project is to understand the effects of egg-parasitoids and predators on stink bugs in New Jersey agriculture. We conducted three sub-projects associated with this grant, and each had specific objectives.
First, we conducted a pilot study in southern New Jersey. The objectives of this study were to:
- Determine how egg parasitism and predation for three stink bug species differs between species and in three different habitats: peach-dominated, soybean-dominated, and forest.
- Compare abundance and diversity of the egg parasitoids associated with different stink bug species in different habitats.
Second, we conducted a comprehensive, landscape-level study in northern New Jersey. The objectives of this study were to:
- Determine how land-use at different scales predict abundances of two stink bug species—the invasive pest Halyomorpha halys and the native predator Podisus maculiventris—in peach orchards.
- Determine how land-use at different scales predicts egg mortality due to parasitoids and predators for two stink bug hosts—H. halys and P. maculiventris—in peach orchards.
- Evaluate reproductive and nonreproductive parasitism by native and nonnative parasitoid wasps utilizing two stink bugs—H. halys and P. maculiventris—as hosts through the application of molecular forensic techniques.
- Monitor spread and establishment of Trissolcus japonicus—a non-native parasitoid wasp that has co-evolved with H. halys and was first detected in in 2017—in New Jersey.
Lastly, we conducted an extension-focused project on the pest status of H. halys in New Jersey. The objectives for this project were to:
- Determine how populations of halys have changed on New Jersey orchards after seven years of sampling by Rutgers IPM Program scouts.
- Determine how the perception of halys as a pest has changed over time among Northeastern tree fruit growers.
Landscape agricultural ecology, or the study of how landscapes surrounding farms influence abundances, diversity, and impact of arthropod pests and natural enemies, has emerged as its own subfield since the turn of the century. Many studies have measured greater control of agricultural pests by natural enemies in structurally complex landscapes, which are often defined as landscapes with a high proportion of natural or semi-natural land or by the diversity of different landscape types in a certain surrounding radius. Mechanistically, natural land may be preferred for natural enemies because of availability of nectar, alternative prey/hosts, and overwintering habitat, and the absence of insecticide sprays and other management. What follows is the idea that conservation of natural land surrounding agriculturally managed land is a win-win for both agricultural producers and conservations (Karp et al., 2018).
To study the effects of natural enemies in a measurable way, studies have deployed sentinel eggs or prey in similar crop blocks surrounded by different landscapes and compared parasitism and predation across blocks. For example, in a study on 26 soybean farm sites in the Midwest, soybean aphid counts were monitored on plants in natural enemy exclusion cages and on unexcluded plants to measure the effect of parasitoids and predators on aphid population growth. In this study, greater landscape diversity predicted greater population control by natural enemies of soybean, and presence of lady beetle predators was correlated with the proportion of forest and grassland in the surrounding landscape (Gardiner et al., 2009). Greater proportion of natural lands in surrounding landscapes also predicted greater parasitism and hyper-parasitism rates of aphids in a study in 18 winter wheat fields in Germany surrounded by different landscapes (Gagic et al., 2011). Similarly, in six Michigan corn fields, parasitism rates of armyworm larvae were higher (13.1% vs. 2.4%) in complex landscapes characterized by smaller crop blocks and higher proportions of hedgerows and wooded habitat (Marino & Landis, 1996).
In addition to pest control by natural enemies, studies have investigated natural enemy abundances and diversity in response to landscape variables. Interestingly, enemy abundance and diversity are not necessarily correlated to pest control services. In the winter wheat field study, aphid-parasitoid food webs were less complex—measured by food web metrics, like mean number of associated host or parasitoid species for all species—in complex landscapes despite greater total parasitism in these systems. This was due to one particular parasitoid species dominating in complex landscapes (Gagic et al., 2011). In the Michigan study, the assemblage of parasitoid species associated with armyworm were similar (4 in simple and 5 in complex landscapes), and again, one parasitoid dominated in complex landscapes (Marino & Landis, 1996).
Clearly, greater landscape diversity and complexity can increase top-down ecological pressure by natural enemies on pests in some systems. However, this phenomenon is not entirely generalizable. In a quantitative synthesis of 46 landscape-level studies, both natural enemies and pests showed a positive response to landscape complexity, as measured by percent natural land in the surrounding landscape. The strength of this response was larger for natural enemies than pests, as natural enemy diversity, abundance, and predation/parasitism performance were all positively correlated with landscape complexity, while for pests, diversity increased, abundance decreased, and crop damage had no significant response with landscape complexity. Furthermore, generalist natural enemies responded to landscape complexity to a greater extent at larger spatial scales than specialists (Chaplin-Kramer et al., 2011). A more recent and comprehensive meta-analysis incorporating data from 143 studies around the world found no consistent trend of the effect of the proportion of natural nor agricultural land in the surrounding landscape on pest or enemy abundance, enemy activity (as measured by sentinel eggs or prey), crop damage, and crop yields (Karp et al., 2018). This study concluded that future landscape agricultural ecology studies should take temporal dynamics into account, include more measures of landscape diversity than just percent natural land, and include measures of local-farm management into models (Karp et al., 2018).
In this study, I investigated how landscape composition and configuration variables predict abundances of adult and nymphal Halyomorpha halys and Podisus maculiventris, egg mortality of H. halys and P. maculiventris sentinel egg masses, and diversity of the egg-parasitoid assemblages associated with each host species. H. halys is an invasive, generalist pest that feeds on over 150 different plants, including cultivated crops: peach, apple, peppers, tomatoes, grapes, corn, soybean, and hazelnut (Leskey & Nielsen, 2018). It is a particularly damaging pest for Mid-Atlantic tree fruit. For instance, during the first major outbreak in 2010, some peach growers saw >90% losses of their crop. Conversely, P. maculiventris is a native, generalist predatory that feeds on over 90 insects, including major pests, like corn earworm and H. halys at the nymphal stage (Marston et al., 1978). Both species are generalists, which poses a unique uncertainty in predicting which landscape variables these insects’ abundances will correlate with, as they both can find food in many habitats.
There are two published studies that have investigated the effect of landscape on H. halys abundance and damages, and none that have examined landscape effects and P. maculiventris. One study of 30 Mid-Atlantic tomato fields concluded that forest edges and number of forest patches correlate positively with H. halys damage, although this study was restricted to 250-m radius around blocks (Rice et al., 2017). Furthermore, an Italian study recorded abundances of H. halys adults and nymphs in pheromone-baited traps on 113 fruit orchards. No effect of landscape on the abundance of adult H. halys was detected, but there was a significant positive effect of percent semi-natural and percent vineyard habitats at a 3 km scale on abundances of nymphs (Tamburini et al., 2023). No studies have looked at landscape composition and configuration effects on sentinel egg mortality of either stink bug despite the high number of sentinel egg mass studies in this system. I predict that landscape variables will impact parasitism greatly, as closely parasitoids that are closely related with pentatomids show strong habitat associations. For example, Telenomus podisi (Hymenoptera: Scelionidae) is a major parasitoid of P. maculiventris and is highly associated with field/vegetable crop habitats, while parasitoids in the genus Trissolcus often parasitize both stink bugs and are associated with forested habitats (Abram et al., 2017).
An egg parasitoid of particular interest in this study is Trissolcus japonicus (Hymenoptera: Scelionidae). T. japonicus is a non-native parasitoid that is closely associated with H. halys in their shared native range in east Asia (Yang et al., 2009). It was first detected in the US in 2015 and has since colonized much of the invaded range of H. halys in North America and Europe (Talamas et al., 2015). Generally, T. japonicus persists in the Mid-Atlantic at low abundances and parasitizes low numbers of H. halys sentinel eggs, so detections of this parasitoid and insights into its landscape ecology is of particular concern (Bergh et al., 2023; Herlihy et al., 2016; Kaser et al., 2018). Furthermore, instances of T. japonicus parasitism of the native beneficial P. maculiventris was of particular interest in this study, as a field study in the Pacific Northwest detected high rates of both reproductive parasitism (24.0%) and nonreproductive parasitism (37.6%) of this host by T. japonicus (Hepler et al., 2020).
In this study, both reproductive parasitism—when a parasitoid wasp oviposits in a host egg and, ultimately, an adult parasitoid successfully hatches—and nonreproductive parasitism—when a developing stink bug dies after a parasitoid oviposits into the egg but no adult parasitoid hatches out—was monitored through the sentinel egg masses. Nonreproductive parasitism is historically understudied because it is difficult to detect, but studies have shown that it happens often in Pentatomid-parasitoid relationships, especially when the species are not co-evolved (Abram et al., 2014, 2016, 2017; Haye et al., 2015). The development of molecular tools has made it possible to detect nonreproductive parasitism by the most common egg-parasitoids associated with Pentatomids in the family Scelionidae and genus Anastatus (Gariepy et al., 2019; Hepler et al., 2020; Stahl et al., 2019). This study system is unique, as there are both native and non-native players at both trophic levels readily in the environment.
Cooperators
- (Researcher)
- (Researcher)
- (Researcher)
- (Researcher)
- (Researcher)
Research
Pilot Study
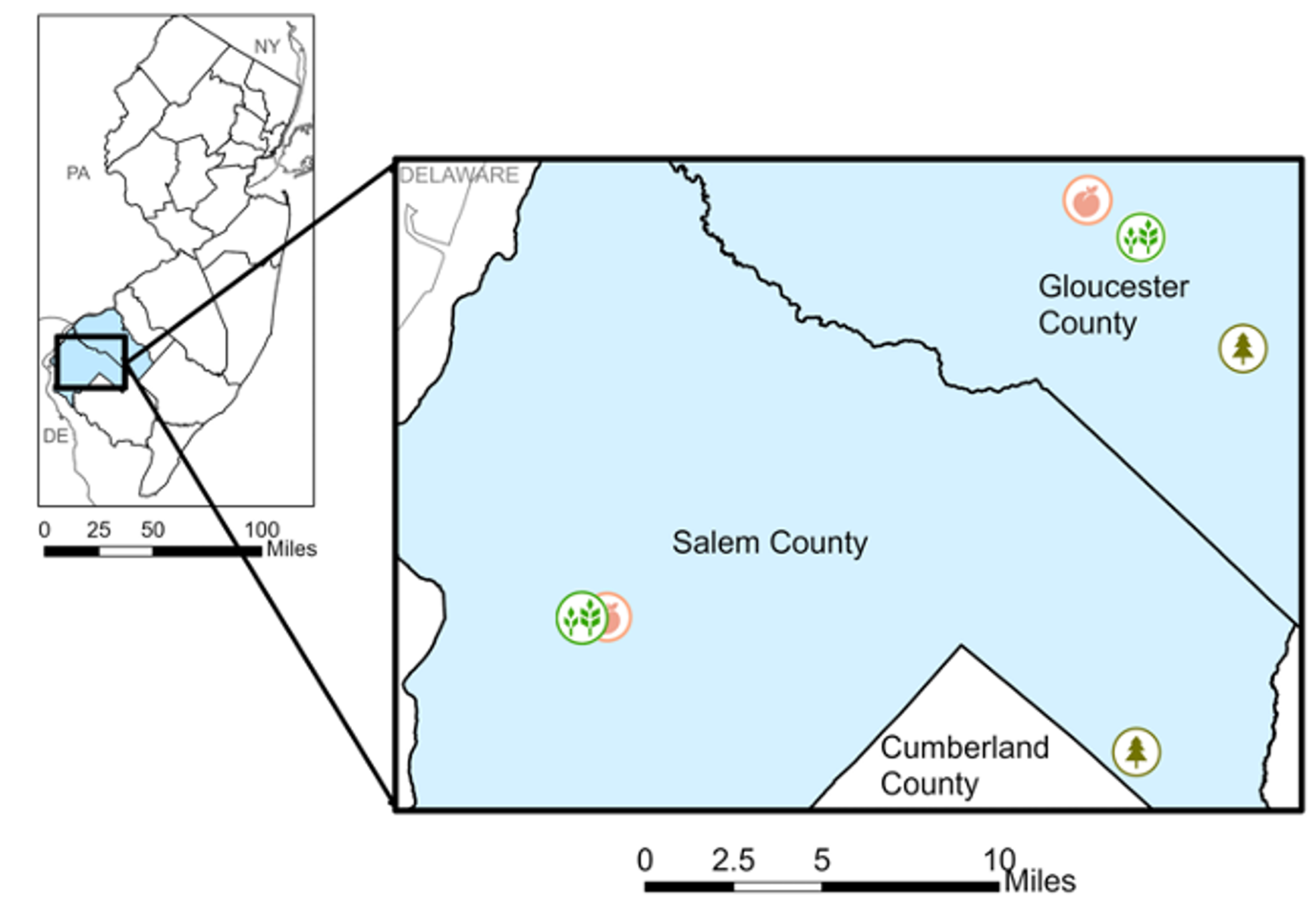
We collected data over two weeks in late July to early August 2022 from two peach, two soybean, and two forested habitats in southern New Jersey (Figure 1). The peach and soybean sites were on operational farms, while one forested site was in Parvin State Park and one was in Glassboro Wildlife Management Area.
Figure 1. Map of study sites for pilot study.
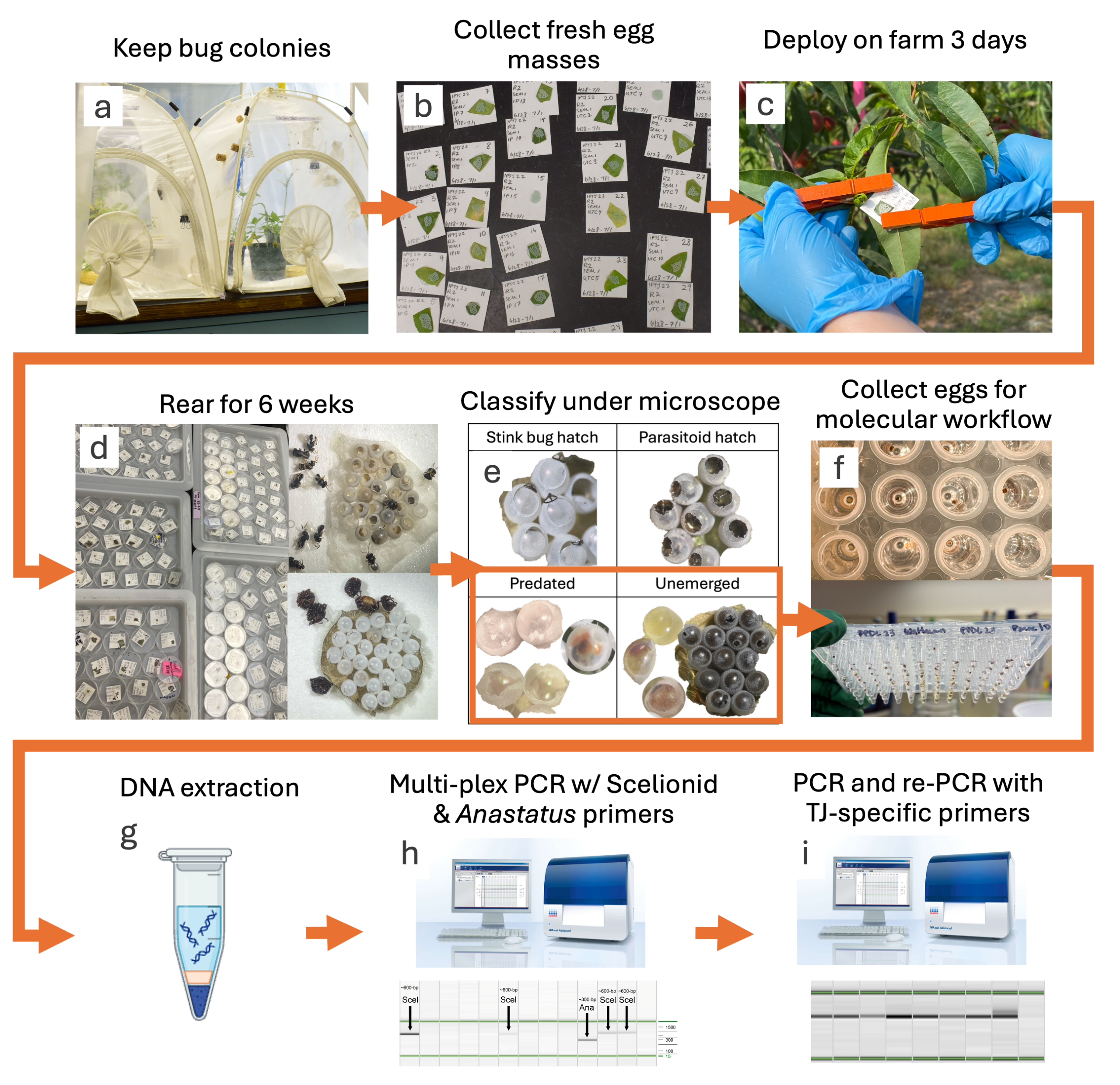
We kept three species of stink bug—Halyomorpha halys, an invasive pest, Euschistus servus, a native pest, and Podisus maculiventris, a native beneficial predator stink bug– in colonies at Rutgers Agricultural Research and Extension Center. The H. halys colony originated from egg masses and adults purchased from the New Jersey Department of Agriculture Phillip Alampi Beneficial Insect Rearing Laboratory. The P. maculiventris colony was a mixture of egg masses and adult insects provided by two USDA-ARS centers, the Beltsville Agricultural Research Center in Maryland and the Beneficial Insects Introduction Research Unit in Delaware. The E. servus colony was started with wild-caught bugs only. We maintained insects in W60 x D60 x H60 cm ‘BugDorms’ (BioQuip) at ~25dC, 60-80% RH, and 16:8 L:D. We reared H. halys and E. servus following modified methods from Abrams et al., 2021. We provided these colonies with cowpea and sunflower plants, water wicks, organic carrots, sweet corn, pumpkin seeds and sesame seeds, and drawer liner and paper towels as oviposition substrates. We provided P. maculiventris cowpea and sunflower plants, the same oviposition substrates, and wax worm and mealworm pupae.
We acquired egg masses from each host from colony tents with a dime-size area of the oviposition substrate and attached them with double-sided tape to 3.5 x 3.5 cm cardstock paper (Figure 2a, b). All egg masses were fresh or were collected fresh and then kept at 8 degrees C for 1-3 days before deployment. At each sample location, we placed eggs both internally (surrounded by at least 30 meters of contiguous habtiat) and externally (at a habitat edge). We attached eggs to the back of peach, soybean, or a known deciduous host plant’s leaves (Figure 2c). We collected egg masses after 3 days in the field and then reared out all stink bugs and parasitoids from masses placed in tight-fitting petri dishes in incubators kept at 25C (Figure 2d). After six weeks, we froze eggs at -20 C for later egg dissection and egg fate determination.
Figure 2. Workflow of stink bug egg handling methods in pilot study.

Egg fates included stink bug nymph emergence, parasitoid emergence, chewing predation, sucking predation, unemerged parasitiod, unemerged nymph, and unemerged unknown (Figure 2e). We identified the unemerged parasitoids and unemerged stink bug nymphs through dissections (Figure 2f). Furthermore, we grouped parasitoid species into morpho-groups to determine which host species is associated with the greatest diversity of parasitoids.
These fates were further classified into egg mortality due to natural enemies (parasitized/predated) and other. I compared egg fate by habitat and species using binomial general linear mixed methods in R Studio.
Overall, we deployed 26 H. halys egg masses (11 in peach, 13 in soybean, and 12 in forest), 16 E. servus egg masses (7 in peach, 4 in soybean, and 5 in forest), and 13 P. maculiventris egg masses (5 in peach, 4 in soybean, and 4 in forest).
Comprehensive, landscape-scale study
Farm selection and study dates
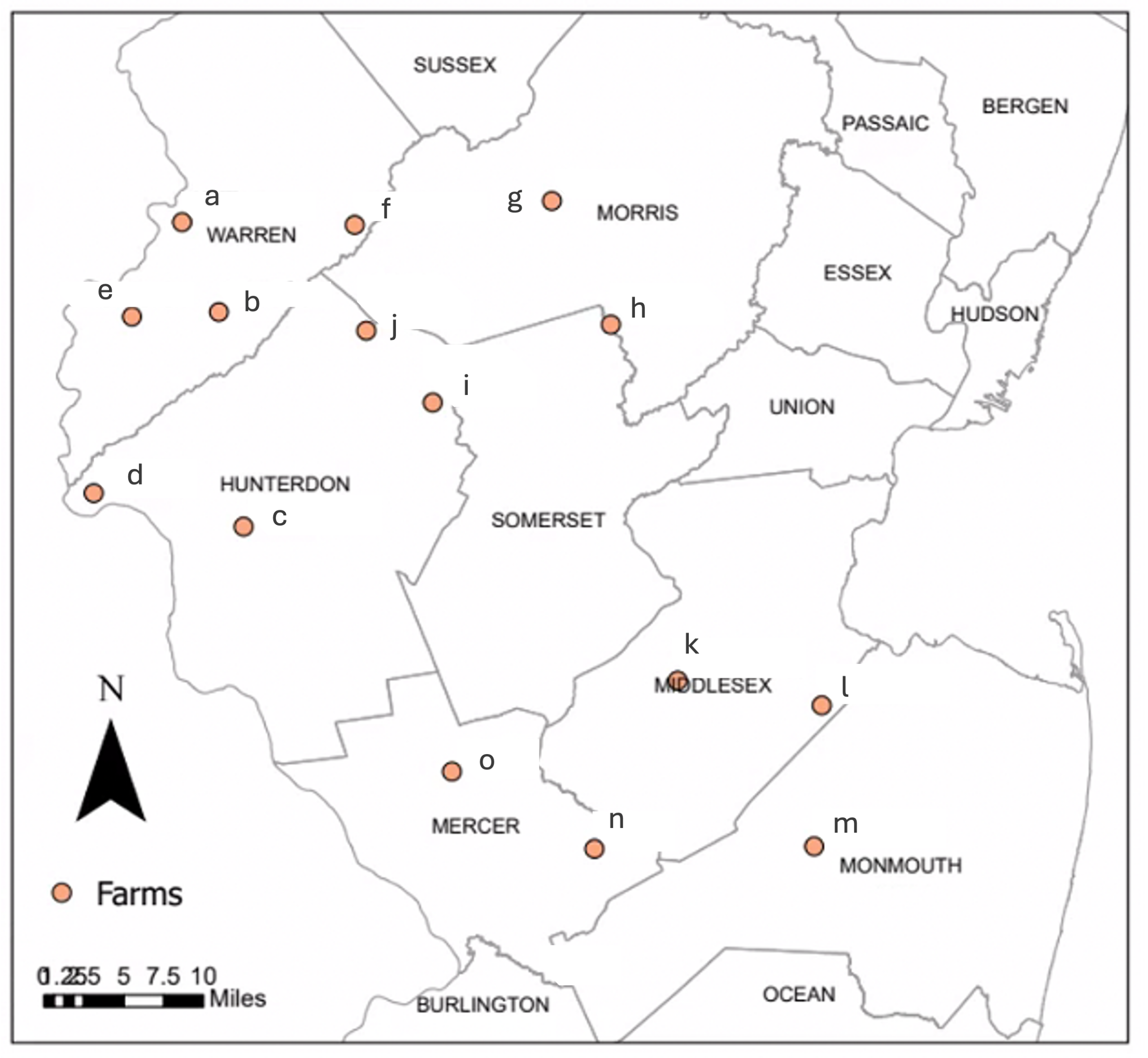
We collected field data from June 11-August 25, 2023 in peach orchard blocks on 15 operational farms in central and northern New Jersey (Figure 3). The closest two sites are ~9 km apart, and the furthest two are ~90 km apart. Peach blocks were between 0.6-3.2 hectares and had at least one wooded or residential edge. We blocked sites into three groups of five that were sampled on the same schedule (farms a-e, f-j, and k-o as labelled in Figure 3). We visited two groups of farms each week, so data from each farm was recorded on 7 out of 11 experimental weeks. Growers from 13 out of 15 farms activity participated in the Rutgers Fruit Integrated Pest Management Program—which provides management suggestions based on trap count and fruit injury data—during the 2023 season, and the other two farms had previously participated in the program. It follows that the farms follow similar management practices.
Figure 3. Study sites on operational farms in central and northern New Jersey. Farms labelled as letters to protect grower privacy.
Stink bug abundance data
We surveyed abundances of H. halys and P. maculiventris with pheromone-baited traps. We deployed traps as 1.3m wooden stakes with a clear sticky trap panel (Peelable Pherocon BMSB STKY trap, Trece Inc) attached at the top with a large binder clip. Traps were bated with pheromone lures. H. halys pheromone lures were purchased from Tréce Inc. and contain a two-component aggregation pheromone produced by H. halys males plus the pheromone of a different stink bug, Plautia stali. These chemicals synergistically attract adults and nymphal H. halys (Weber et al., 2014). P. maculiventris pheromones were prepared using semiochemicals following methods of Kelly et al., 2014 (Figure 4). These pheromones attract adult P. maculiventris. H. halys lures were not replaced over the course of the experiment as per manufacturer recommendations, while P. maculiventris lures were replaced every 2 weeks. We paced traps approximately 50 meters apart from each other on a wooded, meadow, or suburban edge of the peach orchard blocks. We checked traps every 3-11 days, but trap count numbers were standardized when running statistical analyses. Traps were checked 13-15 times at each farm over the season.
Figure 4. A sample P. maculiventris trap.

Egg mass preparation and deployment
We maintained colonies of H. halys and P. maculiventris at Rutgers University Cook Campus in New Brunswick, New Jersey. We maintained colonies in the same manner as detailed above in the pilot study.
We collected fresh (<24 hr old) egg masses from each host from colony tents with a dime-size area of the oviposition substrate and attached them with double-sided tape to 3.5 x 3.5 cm cardstock paper (Figure 5a,b). We photographed egg masses and counted number of eggs deployed. In the field, we attached egg masses to the underside of peach tree leaves with clothespins at the edge of the peach block (Figure 5c). Bordering habitats included apple blocks (9 farms), forest/shrubland (8 farms), mixed annual vegetables/herbs/cut flowers (6 farms), mowed-grass dominant habitats such as parks and mowed fields (6 farms), and corn (3 farms). We collected egg masses after 3 days and placed them singly in sterile tight-fitting petri dishes. Stink bug nymphs and parasitoids were reared out for 6 weeks after deployment in the same conditions as the colonies before being frozen at -20ºC (Figure 5d). In total, we deployed eggs on each farm site during 7 of the 11 weeks of the experiment. The number of egg masses deployed per farm per week depended on colony outputs and was between 1-5 H. halys egg masses and 1-7 Podisus maculiventris egg masses. There were totals of 14-15 H. halys egg masses deployed per farm and 19-21 P. maculiventris egg masses deployed per farm per week. Some were removed from data analysis because they were not found (n=3) or recovered on the ground (n=5). We included a total of 207 of H. halys and 300 P. maculiventris egg masses in statistical analyses.
Figure 5. Workflow of methods for handling egg mass samples in landscape-scale study
Egg fate assessments and molecular analyses
We dissected egg masses under a microscope using sterilized dissecting needles and tweezers (Figure 5e). We classified the fate of each egg as H. halys hatch, parasitoid hatch, predated, and unknown. We recognized sucking and chewing predation following Morrison et al., 2016. We moved emerged parasitoids to individual wells of PCR plates filled with 20 μl of molecular grade ethanol with sterilized paint brushes. Similarly we moved unemerged and predated eggs to individual wells of PCR plates with 20 μl of molecular grade ethanol (Figure 5f). We included an egg parasitized with T. japonicus in each 96-well plate as a positive control along with an empty well as a negative control. All samples were stored at -20ºC between handling.
To identify nonreproductive parasitism, we molecularly analyzed predated and unemerged eggs through a collaboration with Dr. Tara Gariepy’s lab at the Agriculture and Agri-Food Canada London Research and Development Centre in London, Ontario. We extracted DNA from single eggs following a chelex-based DNA extraction method from Gariepy et al. 2014 (Figure 5g). Then, we performed multiple polymerase chain reactions (PCR) with different primer sets. First, we performed a multi-plex PCR with Scelionid- and Anastatus-specific forward primers (Scel-F1 and Ana-361F) that are both paired with the reverse primer HCO2198 of the universal COI primer set (Folmer et al., 1994; Gariepy et al., 2014; Stahl et al., 2019) (Figure 5h). Methodological details, including reaction volumes and solution and thermal cycling conditions, will be detailed in an upcoming publication (Waltman et al., in prep). We used an automated capillary electrophoresis machine (Qiagen Qiaxcel Advanced) to visualize PCR product to score positives for Scelionid and Anastatus parasitism. Then, we ran PCR and re-PCR with T. japonicus- specific primers following methods published in Hepler et al., 2020 (Figure 5i). At the end of this workflow, we knew if each egg had been parasitized nonreproductively by an Anastatus species, by T. japonicus, or by a native Scelionid parasitoid that is not T. japonicus (for eggs negative for T. japonicus DNA but positive for Scelionid DNA).
Landscape classification
We classified the land surrounding each of the 15 study sites by referencing the publicly available Land Use/ Land Cover of New Jersey 2020 polygon dataset published by the NJ Department of Environmental Protection (NJDEP) in ArcGIS Pro (NJ Dept. of Environmental Protection Bureau of GIS; ESRI). We converted the 66 NJDEP specific land-use and land cover classifications into 5 classifications to better suit this project: 1) agricultural (crops, orchards, nurseries, pasture/hay), 2) natural land (forests, shrublands, and wooded wetlands), 3) mowed grass dominant (parks, cemeteries, low-density suburban, etc.), 4) developed (high-density suburban, urban, facility sites), 5) barren/water. Early in the field season, we determined that the agricultural land-use classification by the NJDEP was not consistently valid, likely because of crop rotations and the similar appearance of, for example, tree fruit and tree nursery operations from aerial imagery.
We decided to independently ground-truth and map the agricultural and pastoral land around the farm sites instead of use the NJDEP classifications. We removed land classified as agricultural/pastoral from a copy of the NJDEP polygon layer and instead made a new layer of agricultural land classified by this research team. This new polygon layer was later joined with the NJDEP-classified layer, as the classifications for all non-agricultural land were valid and were utilized for this project. Additionally, two farms had land within 3km that was in Pennsylvania. In these two cases, we used the publicly available National Land Cover Data 2021 dataset to classify land (United States Geological Survey, 2023).
We classified all agricultural and pastoral land in 3 km radii of the 15 farm sites to ensure classification of agricultural land was to-date and fine-scale. This is an area of about 142 km2. From August 10- September 31, 2023, we classified all agricultural and pastoral land in a 3 km radius of the 15 farm sites on foot, when possible, to the following categories: 1) orchards, 2) small fruit (brambles, grape, strawberry), 3) corn, 4) soybean, 6) other annual crops, 6) pasture/hay. We also used high resolution (0.597 m/pixel) satellite imagery obtained from NearMap to inform classification. We compiled classification data in a point vector file in Google Earth and imported this layer imported into ArcGIS Pro. We reclassified and modified (as needed) 2,000+ polygons classified as agricultural land by NJDEP based on the point vector data in ArcGIS Pro. Using ArcGIS Pro tools, including ‘clip’ and ‘calculate geometry’, we calculated the amount of area surrounding each farm into fourteen categories (detailed in Table 1).
Table 1. Description of land-use categories.
|
Land-use categories |
Description |
|
Corn |
Includes row corn and sweet corn |
|
Grain |
Includes wheat, sorghum, and rye |
|
Mixed annual crops |
Includes all annual fruiting vegetables, cut flowers, herbs, etc. Often in high-diversity plantings. Crops in this category include: brassicas, nightshades, cucurbits, mints, Apiales, buckwheat, cut flowers including zinnias |
|
Small fruit |
Includes blueberry, currants, gooseberries, grapes, brambles, strawberries |
|
Soybean/ alfalfa |
Includes soybean, alfalfa |
|
Tree fruit |
Includes peaches, apples, cherries, pears, plums, apricot, pecan |
|
Tree nursery |
Includes tree nurseries, christmas tree farms |
|
Barren |
Includes dirt patches, construction areas, extractive mining operations, and large impermeable paved areas like parking lots, major roadways, railroads, and utility plants |
|
Forest |
Includes coniferous, deciduous, mixed forests, brush, and shrublands, tree plantations, wooded wetlands, and phragmites patches |
|
Grassland |
Primarily refers to managed grasslands, particularly hay and pasture fields, but also preserved meadows, stormwater basins, undeveloped rights-of-way areas, and very early successional areas |
|
Mowed grass dominant |
Includes regularly mowed lawns, athletic fields, cemeteries, recreational land (parks) |
|
Shrubland |
Includes deciduous scrub/shrublands and wetlands, herbaceous areas with shrubs |
|
Water |
Includes artificial lakes, exposed flats, natural lakes, streams and canals |
|
Suburban |
Includes residential areas that are low to high density; generally dwelling areas that also feature a fair amount of lawns and shade trees |
Statistical analyses
To tackle objectives one and two, we ran principal component analyses (PCAs) using percent of land surrounding each farm that falls into each of the 14 different land-use categories at two different scales: 1.5km and 3 km. A principal component analysis condenses many variables into a few uncorrelated variables to explain the most variance in the data as possible. Similar methods have been utilized in previous agricultural landscape ecology studies (Gardiner et al., 2009; Tamburini et al., 2023). We ran the PCAs and conducted the rest of the statistical analyses in RStudio. We also used the vegan() package in RStudio to calculate the landscape diversity and evenness of the surrounding landscape by calculating the gini-Simpson index using land-use category instead of species as in Gardiner et al. 2009. This index returns a value between 0-1 for the land surrounding each farm where 0 is low diversity and 1 is high land-use diversity.
To determine how land-use predicts abundances of the two stink bugs, we modeled the count of adult bugs in each trap using generalized linear mixed models (glmms) with Poisson distributions. All models included week number and farm as random intercepts and an offset of the log of the number of days between trap-checks. For both 1.5km and 3km, we compared a set of six models with the following fixed effects: the interaction between principal component 1 (PC 1)* species, the interaction between principal component 2 (PC 2)* species, the interaction between gini-Simpson index for land-use diversity* species, and pairs of these two interactions. A model including species interactions with all three landscape variables failed to converge. We selected the best-fit model for each scale by Akaike Information Criterion (AIC), which measures model-fit, and compared all twelve models (six for both 3km and 1.5km radii) to select the best-fit model.
To determine how land-use predicts egg mortality of the two stink bugs, we constructed a series of glmms with binomial distributions where the response variable was the paired number of eggs killed by natural enemies (eggs reproductively parasitized + eggs nonreproductively parasitized + eggs predated) and number of eggs not killed by natural enemies for each egg mass. As with the stink bug trap count models, all models included week number and farm as random effects. We constructed and compared models with land-use variables at 1.5km and 3km as fixed effects in the same manner as with the stink bug abundance models to compare a total of twelve models.
Additionally, we ran an additional model investigating how the local habitat predicts egg mortality for both species. For the local habitat model, we limited the data set to only include egg masses we had deployed on the edge of a peach block and a habitat dominated by one of the four most common border habitats: forest/shrubland (represented on 8 farms), mixed annual crops (6 farms), mowed grass dominant (6 farms), or apples (9 farms). For an egg deployment site to be included, the border habitat type had to be at least the second-most represented habitat type in a 50m radius (only after peach). No other border habitat was represented on more than 4 of the 15 farms. This habitat dataset included 361 egg masses (71.2% of total). We modeled the same response variable by the interaction of habitat type*species with the same random effects as described above. We could not run local habitat models to predict trap count because all traps were placed between peach blocks and forest/shrubland except for three.
For all best-fit models, we extracted model estimates and 95% confidence estimates in R Studio and visualized model predictions with ggplot2().
Extension project
We theorize that H. halys populations and pest prevalence have become reduced because of the arrival of T. japonicus to North America circa 2015. To investigate if numbers in New Jersey have actually gone down and if growers’ perception of the pest has also become reduced, we collaborated with Rutgers University extension agents Janine Spies and Kaitlin Quinn. Spies and Quinn run the Rutgers IPM program and supervise scouts who collect trapping and fruit injury data on farms across the state each week during the growing season.
We consolidated weekly H. halys trap data recorded on 27 operational New Jersey apple and peach orchards in 2018 and 2020-2025. These data are recorded by Rutgers IPM scouts each week to monitor and inform spray decisions. The IPM program utilizes the same pheromone-baited traps with clear sticky cards as we had used in my landscape-level study detailed above. They are placed on the edge of apple or peach blocks bordered by a wooded edge. We visualized this data by graphing average trap count per week per year in RStudio.
Lastly, we investigated how the pest priority of H. halys has changed over time by consolidating survey response data collected and distributed by other extension personnel, namely Janine Spies at Rutgers University and Anna Wallis at Cornell University. For more details about the surveys, please see the Education and Outreach section of this report. Again, we did not distribute or administer these surveys for this project, but examining the responses to H. halys-specific questions helps inform this project
Pilot study:
Egg mortality due to parasitoids and predators was highest in the soybean for all three species of stink bug (Figure 6, 7). Overall, P. maculiventris had the highest egg mortality due to parasitoids and predators, but this was habitat-dependent. For instances, P. maculiventris had 99% egg parasitism in soybean but about 30% in peach (Figure 6, 7). Egg mortality was significantly higher for P. maculiventris than for H. halys but neither had significantly greater egg mortality than E. servus.
Figure 6. Egg fate for each host species in each habitat as percent of total eggs in each fate.
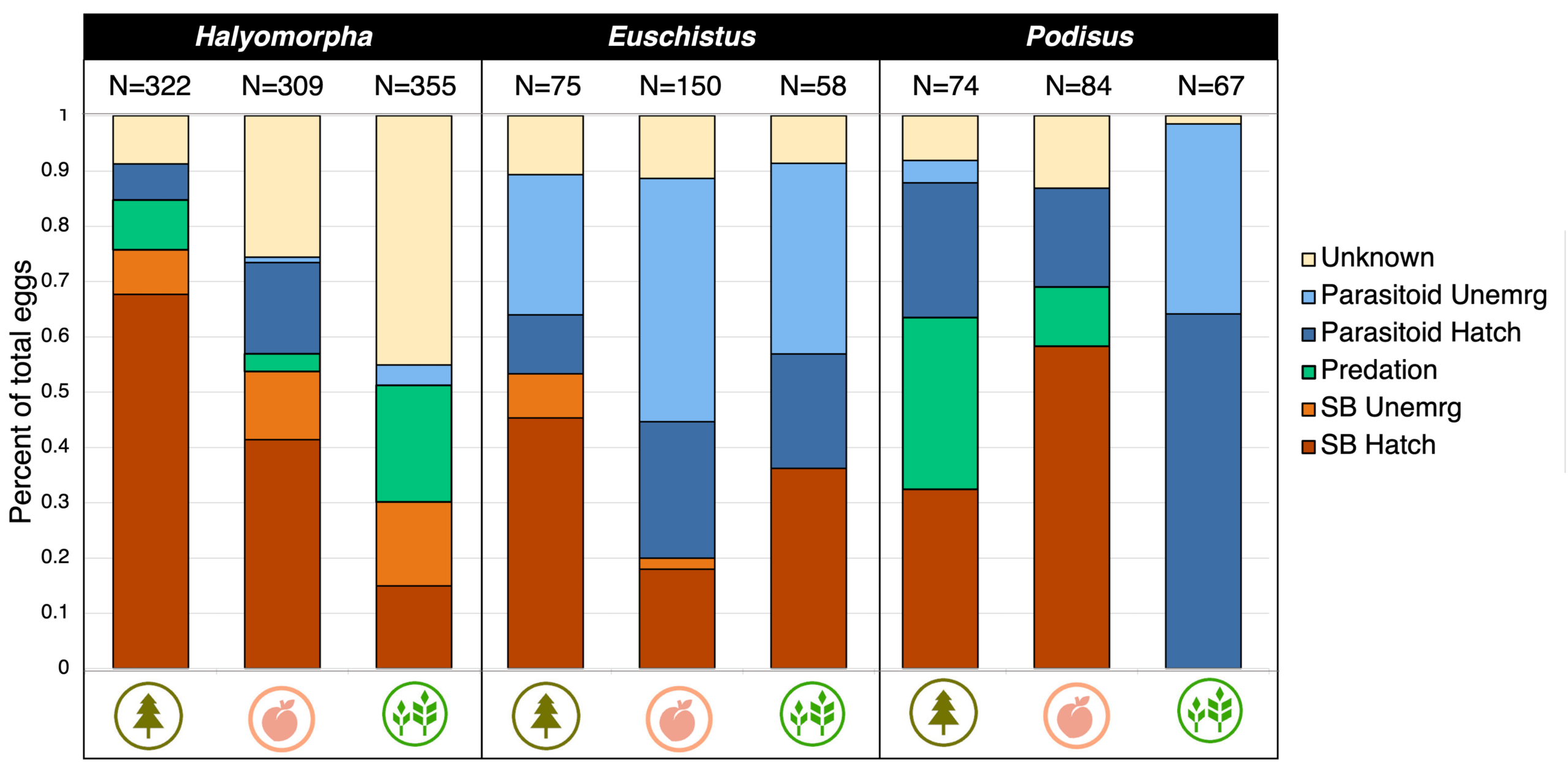
Figure 7. Percent of egg masses with egg mortality due to parasitism or egg predation across species and habitats.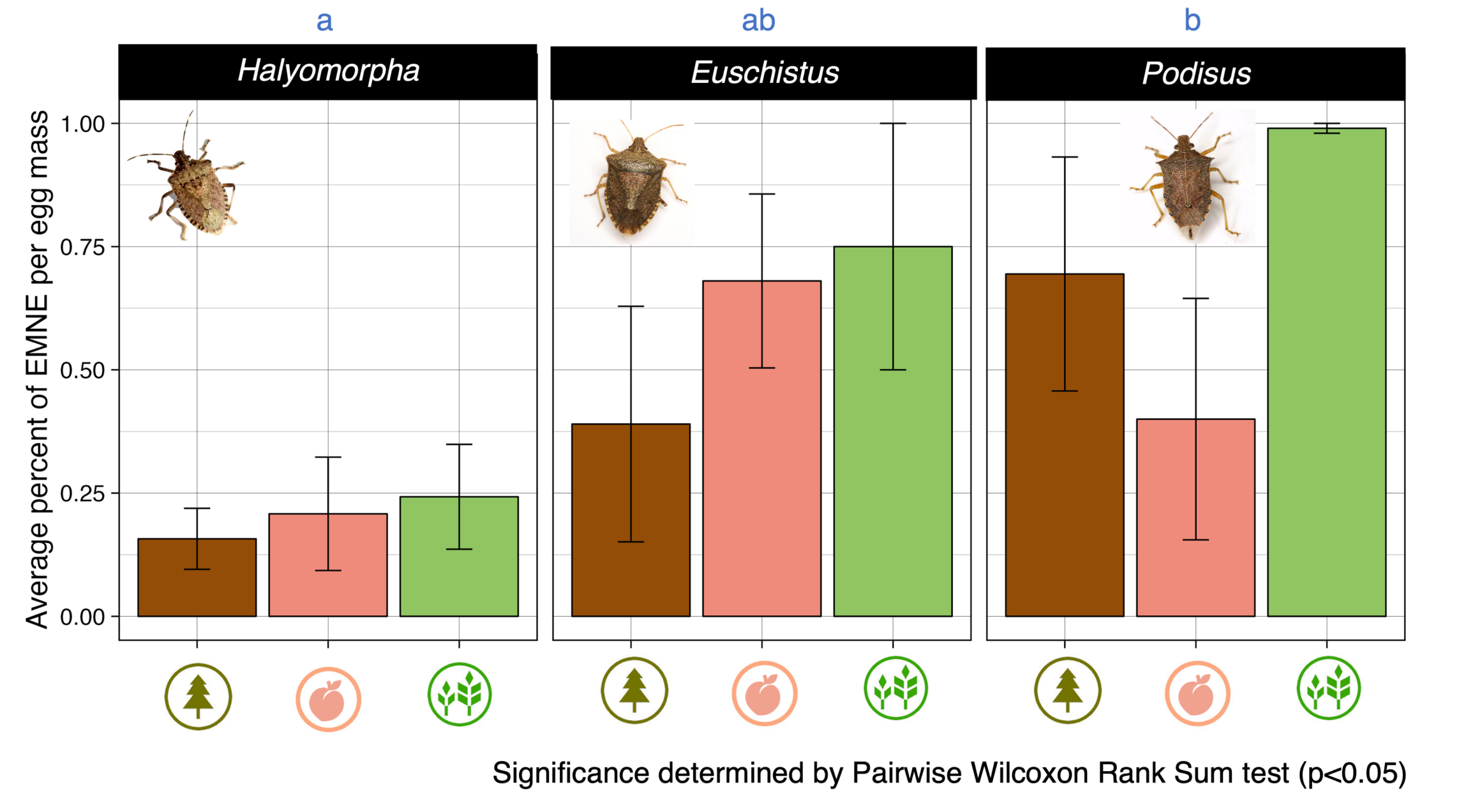
We detected four species of parasitoids with this study. This included T. japonicus, which was detected parasitizing H. halys in peach and forest, and two other, native Scelionid wasps. These included Telenomus podisi, which parasitized all three host species and was most commonly detected in soybean, and an unknown Trissolcus species which parasitized P. maculiventris in forest. Anastatus, a hyper-parasitiod genus that is less host-specific than the others, parasitized H. halys and E. servus and was detected in all three habitats (Figure 8).
Figure 8. Parasitoid species associated with eggs of the three different stink bugs. Dotted line indicates that only nonreproductive parasitism was observed with a particular parasitoid-stink bug pairing.

Comprehensive landscape-level study:
Land-use variables
The PCA of land-use data at 3km radii around farms gave two principal components (PCs) that together described 82% of the data’s variance. Similarly, the first two PCs yielded by the PCA with the 1.5km radii data described 74.2% of the variance. Correlations of PCs with each land-use type were similar for the PCAs at the two scales. PC1 was most positively correlated with forest cover and was most negatively correlated with suburban land-use. PC2 was most positively correlated with suburban land-use and negatively correlated with corn, soybean/alfalfa, and grassland habitat, which includes pasture and hay operations (Figure 9). Land-use variables with small values for PC1 and PC2, such as tree fruit, small fruit, and mixed annual crops, had similar percent landcover around most farms in the study.
Figure 9. Correlations between PCs and land-use variables at two different scales, 3km and 1.5km radii around farm sites.
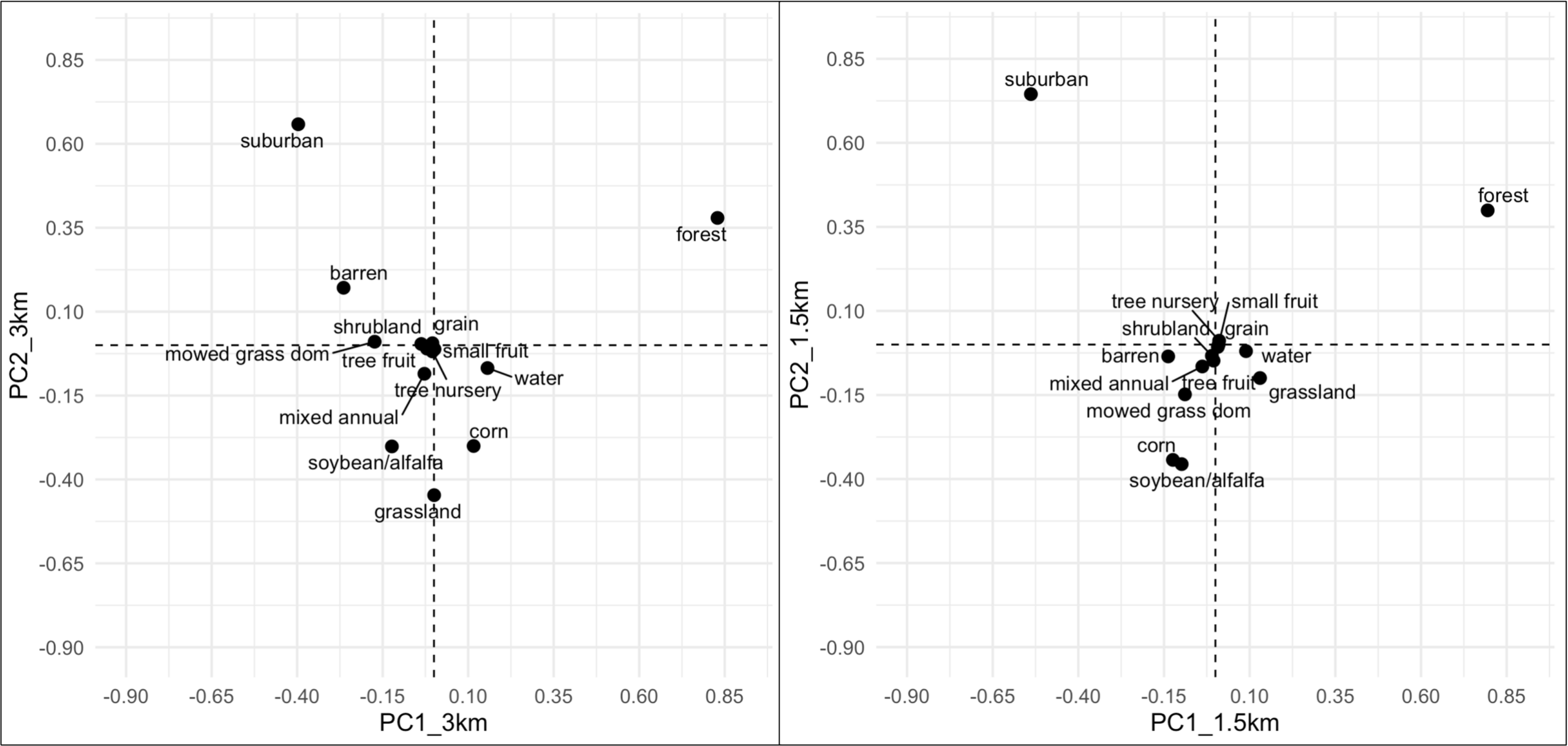
Values for the gini-Simpson (D) value ranged from 0.558-0.8509 at 1.5km radii and 0.6302-0.8463 at 3km radii around farms. Farms typically had similar values for D at both scales.
Stink bug abundances
The average trap count of H. halys adults per week was 0.323 bugs/trap and ranged from 0-3 bugs per week. For P. maculiventris, the average trap capture was 2.842 bugs/trap and ranged from 0-15 bugs per trap per week. These two averages are not necessarily comparable, as the strength of the pheromone bait on the traps was not necessarily equal. Still, the field season when data was collected was a particularly low abundance year for H. halys in the region as reported by growers and extension professionals. This is a limitation of our data set, as H. halys associations with land-use variables may have been more pronounced if our data was not as zero-inflated as it was.
The best-fit model of the twelve models compared that modeled trap count of stink bugs by interactions between species and landscape variables at 1.5km and 3km scales was:
trap count ~ D[3km]*Species + offset(log(days)) + (1| Farm + Week), family=Poisson
where D[3km] is the gini-Simpson index of diversity applied to land-use type at a 3km radius around farms and days is the number of days in between trap counts were recorded. The interaction between D[3km] and Species was significant (X2 = 11.1, df=1, p < 0.001). Principal components of land-use variables were not included in the best-fit model for trap count. The best-fit model predicted a strong negative effect of land-use diversity on Podisus maculiventris trap count and a weak positive effect of diversity on H. halys trap count (Figure 10).
Figure 10. Trap count (per week) of each species as predicted by Simpson’s D at a 3km radius for land-use type is plotted as lines with 95% confidence intervals. The points are data collected during the study; larger points indicate multiple instances where a given trap count was recorded at a given farm.
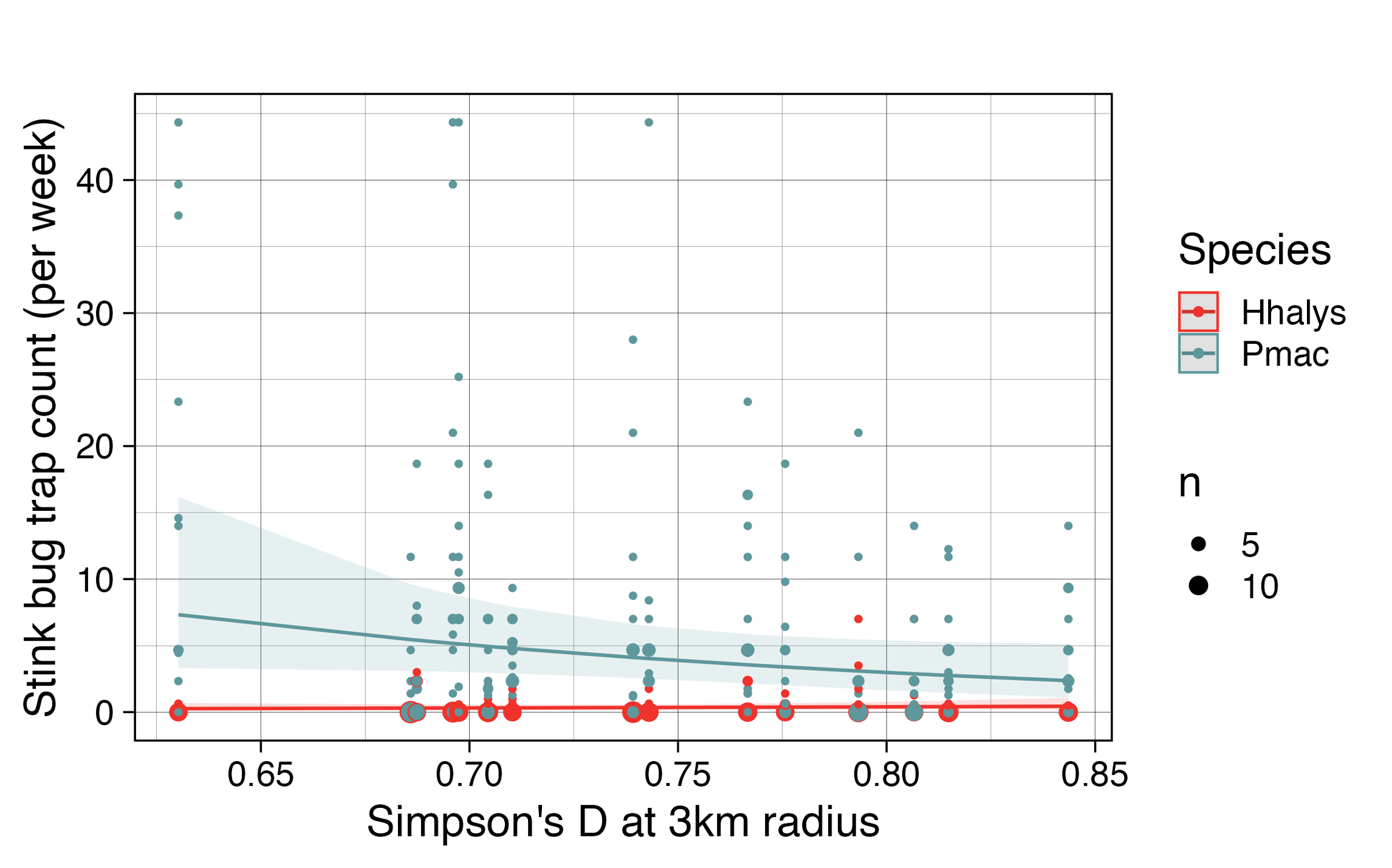
Generally, low-diversity landscapes were dominated by large forest plots. H. halys can utilize arboreal host plants common in forested areas in New Jersey which is likely why it was positively associated with forest. P. maculiventris, on the other hand, often feeds on soft-bodied lepidopteran larvae, which may be easier to find in areas dominated by agriculture and open land-use types as opposed to forests.
Stink bug egg mortality by parasitoids and predators
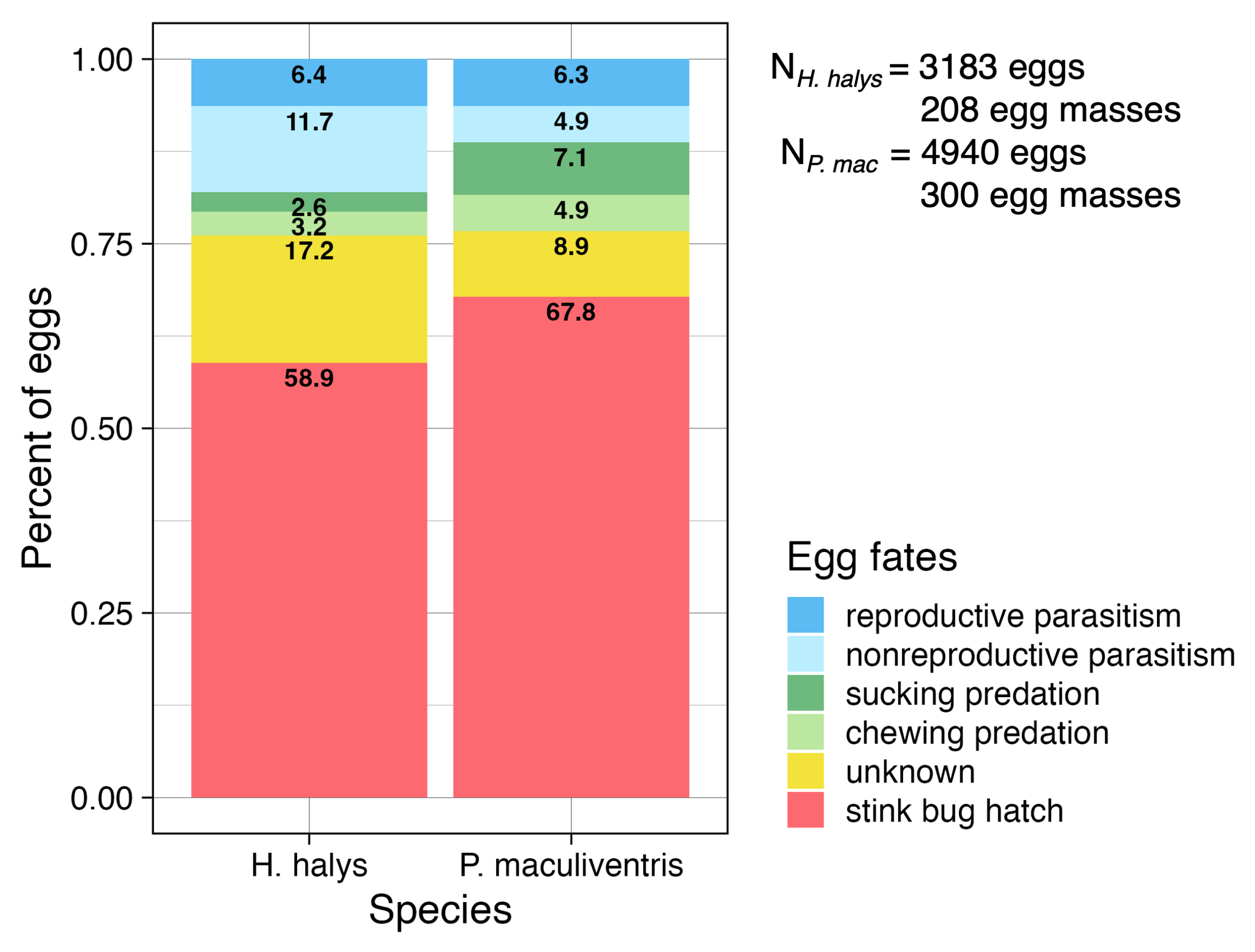
Overall, parasitoids and predators caused nearly equal mortality to H. halys eggs (23.9% ) and P. maculientris eggs (23.2%) (Figure 11). Eggs that did not hatch that were not predated and did not test positively for parasitoid DNA were labelled as ‘unknown’ eggs and accounted for 17.2% of H. halys and 8.9% of P. maculiventris eggs. Eggs in this category could have been unfertilized by stink bug mothers, died due to climatic variables. They also could have been nonreproductively parasitized by parasitoids that were not our taxa of interest and/or parasitized by one of these taxa but did not have enough parasitoid DNA to be detected in our methods. It is likely that some proportion of the unknown eggs were nonreproductively parasitized. These methods will continue to be honed by collaborators in the Gariepy lab in ongoing collaborative studies.
Figure 11. Percent of deployed H. halys and P. maculiventris eggs in each egg fate category.
The best-fit model of the twelve egg mortality models with interactions between species and landscape variables at 1.5km and 3km scales was:
(eggs para/pred, eggs not) ~ PC1[1.5k]*Species + PC2[1.5k]*Species + (1| Farm + Week), family=binomial
The interaction between PC1[1.5km]*Species was significant (X2 = 7.18, df=1, p = 0.007) and PC2[1.5km]*Species was also significant (X2 = 73.40, df=1, p < 0.001). Landscape diversity measured by Simpson’s D was not included in the best-fit model.
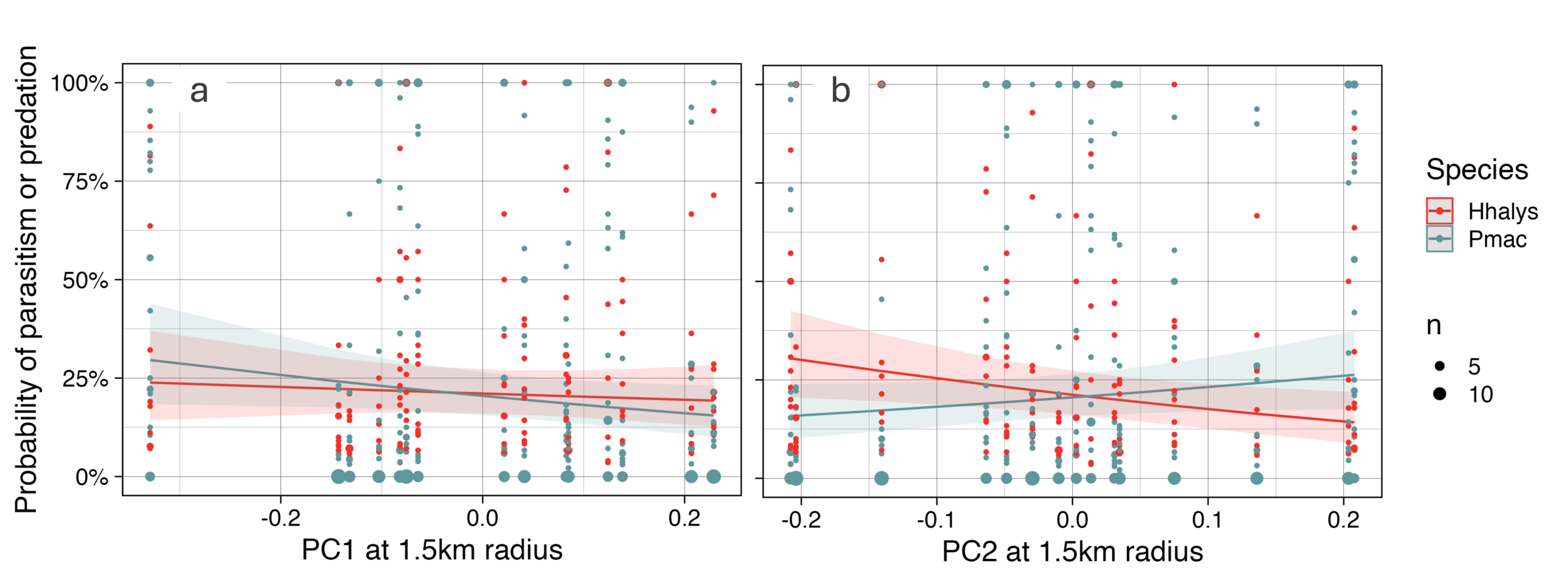
For H. halys, there was almost no correlation between egg mortality by natural enemies and PC1[1.5km] and a negative correlation with PC2[1.5km]. For P. maculiventris, there was a negative correlation with PC1[1.5km] and positive correlation with PC2[1.5] (Figure 12a, b). It is somewhat surprising that biological control of H. halys was not positively correlated with forested land-use in the surrounding landscape. H. halys is typically associated with forested habitats and there are no pesticide sprays in these areas, so one might think natural enemies are abundant there. However, H. halys does overwinter in suburban dwellings, which may explain the positive correlation between natural enemy activity and suburban land-use. It is unsurprising that P. maculiventris’ natural enemy activity is negatively correlated with PC2, which is negatively correlated with soybean, as P. maculiventris commonly occurs in soybean, and soybean is thought to be good habitat for parasitoid wasps due to its extrafloral nectaries.
Figure 12. Probability of egg mortality due to parasitoids and predators of each species as predicted by PC1 and PC2 at a 1.5km radius is plotted as lines with 95% confidence intervals. The points are data collected during the study; larger points indicate multiple instances where a given egg mass at a given farm had a given mortality due to parasitoids and predators.
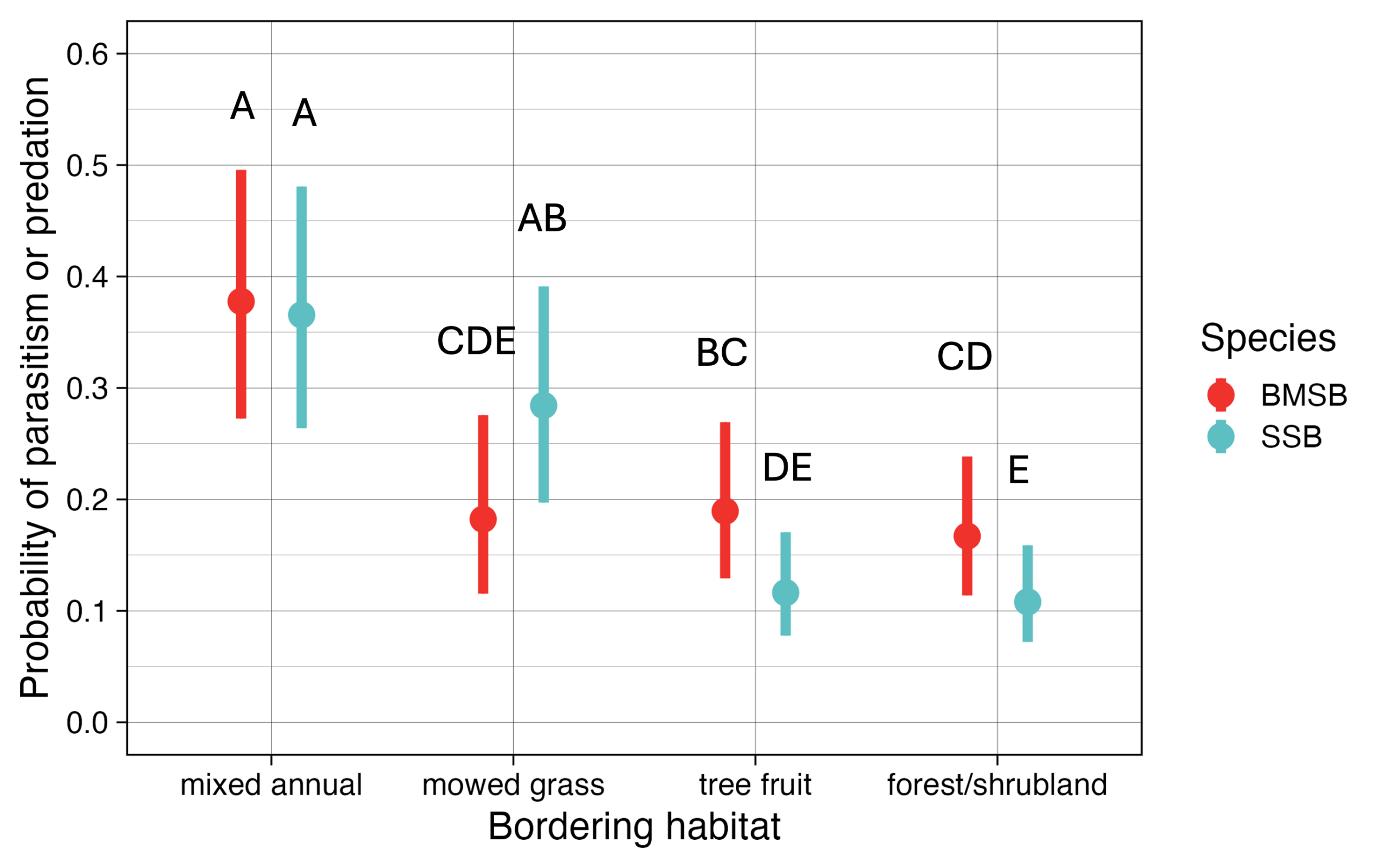
For the habitat analysis, there was a significant interaction between habitat*species (X2 = 33.5, df=3, p < 0.001). Overall, both species were most likely to be parasitized or predated across from mixed annual crops plantings, and in this habitat, there was no significant difference between species (Figure 13). This suggests that growers should consider planting diverse plantings of mixed fruiting vegetables, herbs, or cut flowers near orchards to maximize biological control of H. halys. Furthermore, P. maculiventris was significantly more likely to be parasitized/predated than H. halys across from mowed grass habitats, and H. halys was significantly more likely to be parasitized/predated than P. maculiventris across from tree fruit (apples/cherries) and forest/shrubland habitats (Figure 13). This makes sense, as P. maculiventris is typically associated with grassy habitats, while H. halys is typically associated with orchards and forested areas.
Figure 13. Probability of parasitism or predation of eggs of H. halys and P. maculiventris in border row peaches across from four habitats. The figure shows mean and 95% confidence intervals as predicted by the model. Letters indicate statistical significance (p < 0.05, tukey method corrected for eight treatments).
Egg parasitism and predation details and status of T. japonicus
Halyomorpha halys and P. maculiventris eggs were reproductively parasitized to nearly equal proportions (6.4% of H. halys eggs and 6.3% of P. maculiventris eggs) (Figure 11). However, H. halys eggs were nonreproductively parasitized more than twice as often as P. maculiventris eggs (11.7% vs 4.9% of eggs) as detected by the molecular methods (Figure 11). The greater total parasitism rates for P. maculiventris was not expected, as H. halys is invasive and has only been present in NJ for approximately 30 years, while P. maculiventris is native and has co-evolved with native parasitoids for longer. However, H. halys is more generally associated with peach habitats and P. maculiventris is not, so parasitoids that prefer P. maculiventris may be less likely to seek eggs as reproductive resources in peach orchards. Additionally, T. japonicus was detected in this study and consistently reproductively and nonreproductively parasitized H. halys (5.62% and 2.80% of eggs, respectively) more than P.maculiventris (0% and 1.05% of eggs, respectively; Table 2).
Table 2. Percent of total eggs for each host species reproductively and nonreproductively parasitized by the three parasitoid taxa. Nonreproductively parasitized eggs that tested positive for DNA of an Anastatus parasitoid and either TJ or non-TJ Scelionid were counted toward totals for both parasitoid taxa.
|
Host species |
Parasitoid taxa |
Reproductive parasitism (% total eggs) |
Nonreproductive parasitism (% total eggs) |
Total mortality caused to eggs (% total eggs) |
|
H. halys |
Trissolcus japonicus |
5.62 |
2.80 |
8.42 |
|
Non-T. japonicus Scelinoids |
0.31 |
8.39 |
8.70 |
|
|
Anastatus spp |
0.41 |
0.60 |
1.01 |
|
|
P. maculiventris |
Trissolcus japonicus |
0 |
1.05 |
1.05 |
|
Non-T. japonicus Scelinoids |
5.98 |
0.83 |
6.81 |
|
|
Anastatus spp |
0 |
3.14 |
3.14 |
Interestingly, a similar proportion of the invasive H. halys eggs were parasitized (reproductively or nonreproductively) by T. japonicus, a non-native parasitoid (8.42%) and native parasitoids (9.71%) (Table 2). Most T. japonicus parasitism was reproductive, while most non-native parasitism was non-reproductive. This was expected given the literature. Still, the amount of biological control of H. halys by native parasitoids is historically under-reported and not considered due to the cryptic nature of nonreproductive parasitism.
Eggs with detected non-reproductive parasitism by Anastatus species, which is known to hyper-parasitize eggs and can hyper-parasitize eggs first parasitized by Scelionids, were sometimes also positive for DNA of T. japonicus (4.0% of Anastatus non-reproductive parasitism events) and were sometimes also positive for DNA of a non-T. japonicus Scelionid (0.6% of these events). It is possible that the arrival and spread of T. japonicus is providing opportunity for Anastatus to utilize stink bug hosts more than parasitoids in this genus were before its arrival.
Trissolcus japonicus was detected on all fifteen farms in the study. This is promising. Although it only parasitized 8.42% of H. halys eggs—less than what is reported in China where both are native—this is larger than previously published parasitism rates by T. japonicus in New Jersey orchards, which were 1.4% in 2017 (Kaser et al. 2018). Additionally, the non-target impact of T. japonicus on P. maculiventris was small, with only 1.05% of eggs parasitized (Table 2). This suggests that releasing T. japonicus on farms would not have a significant negative impact on populations of P. maculiventris.
A higher proportion of P. maculiventris eggs were predated by sucking and chewing predators (7.1% and 4.9%, respectively) than H. halys eggs (2.6% and 3.2%). Common predators found in orchards that may have contributed to this predation include meadow katydids, jumping spiders, pirate bugs, and lacewings (Morrison III et al. 2016).
Extension project
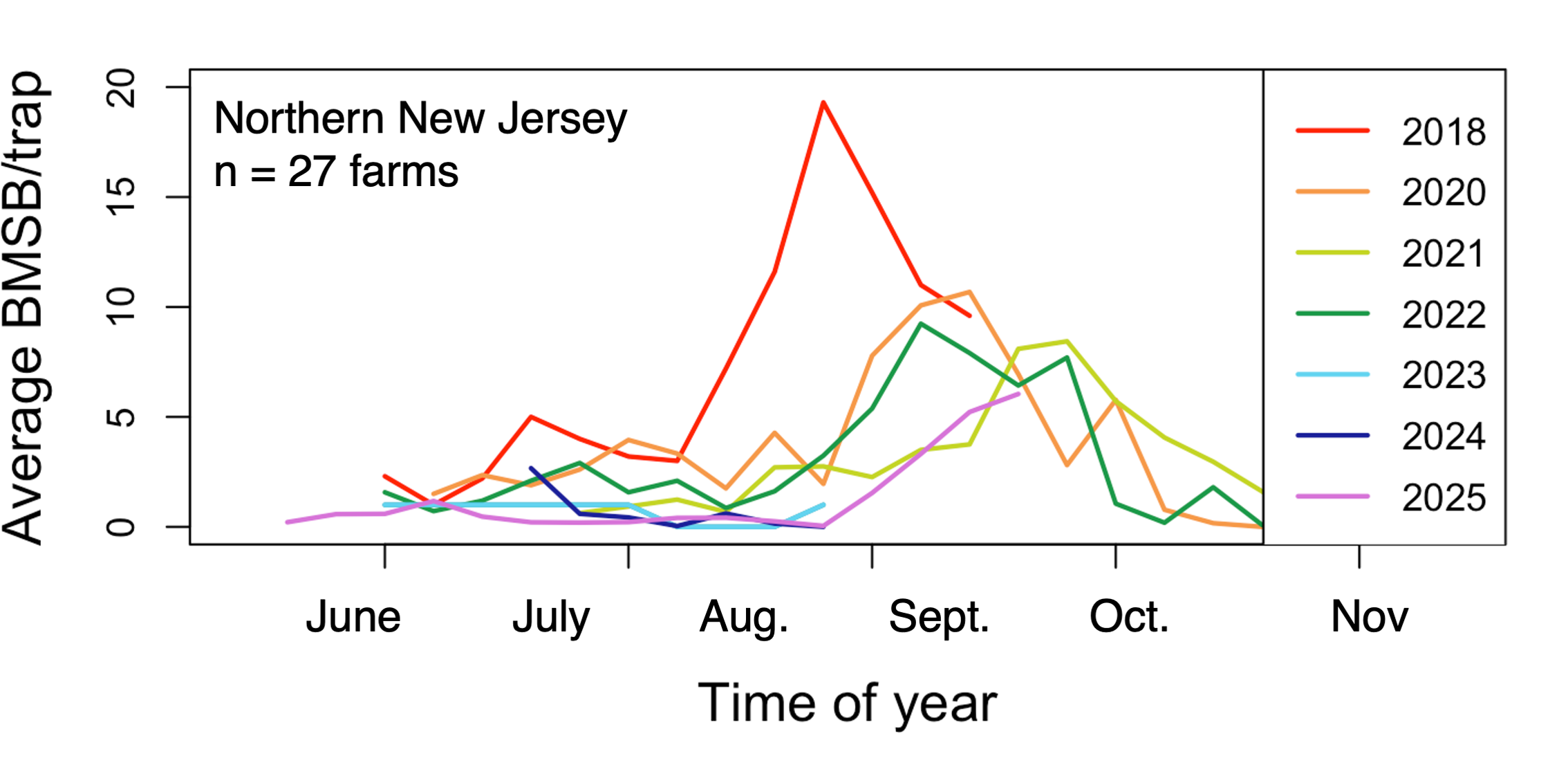
Generally, populations of H. halys have become lower over time. The largest population peek for H. halys during the study period when these data were collected was in 2018 (Figure 13). In 2020-2022, the population is lower and is fairly consistent year-to-year. Unfortunately, there are gaps in the data in 2023 and 2024, as data was only collected during the summer months and not during September, when H. halys populations tend to peak. Still, trap counts during July and August were generally low in 2023 and 2024. The apparent trend in 2025 also suggests a lower population compared to 2020-2022 (Figure 13). While the data presented in the figure only span seven years, which is not incredibly long for studying long-term population trends for insects, these data illustrate a trend over time on operational farms in the Rutgers IPM program.
Figure 13. Average weekly H. halys trap counts per week over 7 years. All data collected on 27 Northern New Jersey peach and apple farms by Rutgers IPM Program scouts.
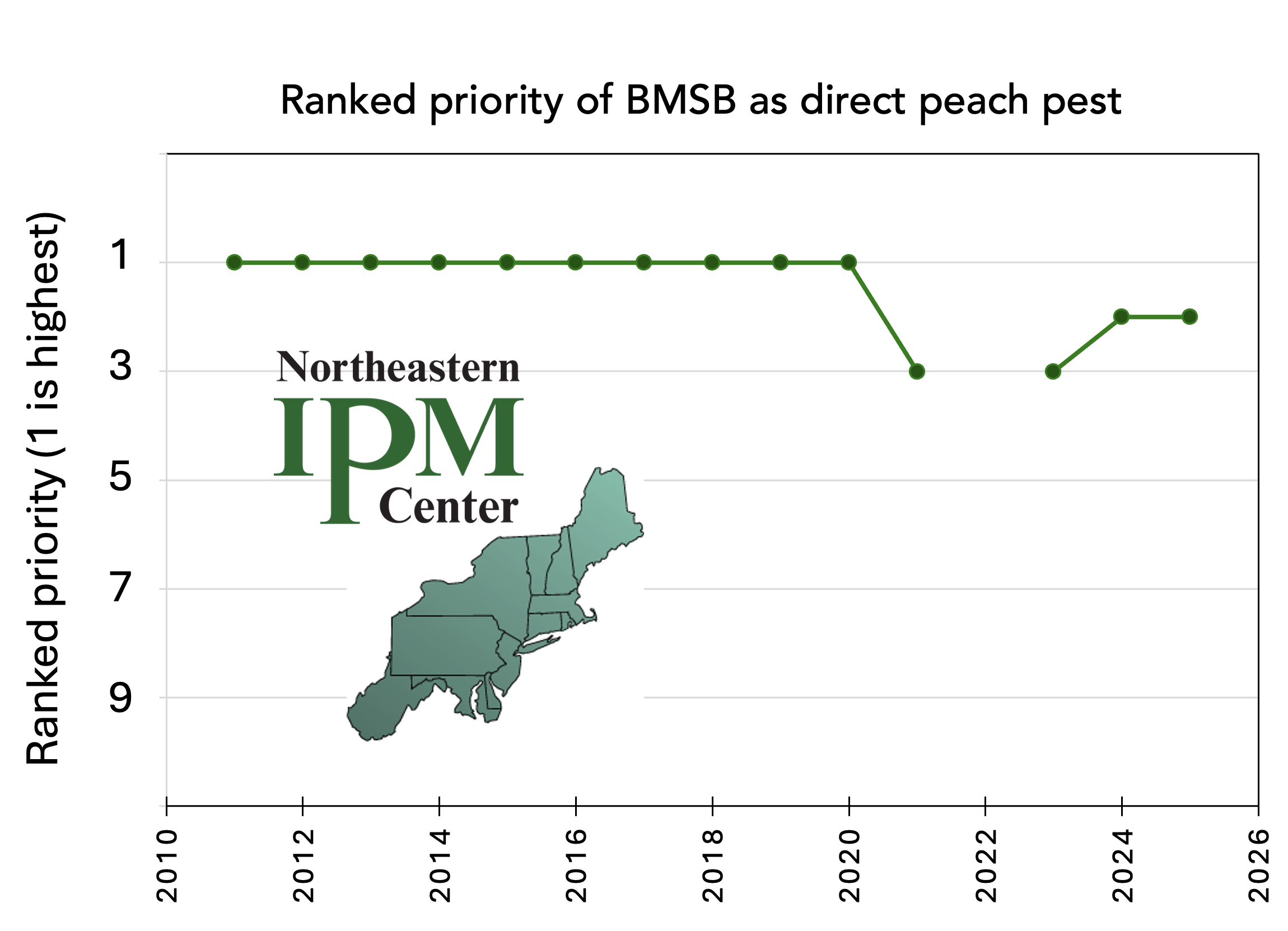
Over time, H. halys has remained a moderate and persistent pest, yet it has become less of a high priority pest in recent years. The Northeast Tree Fruit IPM Working Group (NETFIPM) members consistently indicated H. halys as the highest priority pest direct pest of peach from 2011-2020; however, it dropped to number three in 2021, was number three again in 2023, and was number two in 2024 and 2025 (Anna Wallis via Northeast IPM Center website) (Figure 14). Furthermore, in a 2024 grower survey of 29 apple and peach growers in New Jersey, 21% of respondents described H. halys as a significant issue, 38% occasional issue, 17% as no issue, and 24% did not respond (distributed and communicated by Janine Spies, 2024). In this survey, H. halys was the second-most identified insect pest ranked as significant or occasional issue. Despite the apparent reduction in population size and change in priority-ranking, it is clear that H. halys continues to cause pest problems for a majority of NJ tree fruit growers.
Figure 14. Responses of the Northeast Tree Fruit IPM Working Group (consultants and extension personnel) when asked to rank the priority of direct peach pests in the northeast. 1 indicates highest priority (Wallis, 2011-2025 via Northeastern IPM center).
The overarching goal of this project was to understand how egg‑parasitoids and predators influence populations of the invasive pest Halyomorpha halys and the native predator Podisus maculiventris in New Jersey. Across three sub‑projects, I quantified natural enemy impacts, determined how habitat-scale and landscape-scale composition affects multi-trophic interactions between stink bugs and their natural enemies, and evaluated long‑term trends in H. halys populations and grower perceptions of the pest.
This pilot study monitored egg predation and parasitism of H. halys, P. maculiventris, and Euschistus servus, a native pest, in peach, soybean, and forested habitats in southern New Jersey in 2022. Although sampling effort was limited, trends emerged. For all species, egg mortality due to natural enemies was highest in soybean. Furthermore, P. maculiventris had significantly greater egg mortality than H. halys, although this was only significant in forested and soybean habitats, not peach. Trissolcus japonicus, H. halys’ closely co-evolved egg parasitoid from Asia, parasitized H. halys, not the native stink bugs, in forest and peach habitats, while Telenomus podisi, which is closely co-evolved with P. maculiventris in the US, parasitized all three species and was most-commonly found in soybean. This preliminary work solidified egg mass handling methods and illuminated the value of a larger-scale landscape study to investigate how habitat preferences of stink bugs and their natural enemies scale up to predict biological control on farms.
In the comprehensive landscape‑level study, I monitored H. halys and P. maculiventris populations and egg mortality in northern New Jersey in 2023. Over 15 farms, we deployed pheromone-baited traps and 507 egg masses over 11 weeks. In addition to identifying reproductive parasitism and predation, we applied a novel molecular workflow to identify nonreproductive parasitism by multiple parasitoid taxa. We performed landscape analyses at two scales and included a measure of land-use diversity and evenness and a Principal Component Analysis to reduce many land-use categories into two, uncorrelated variables. H. halys trap captures increased slightly with greater land‑use diversity, likely reflecting the species’ ability to exploit many host plants, including deciduous trees and agricultural commodities. In contrast, P. maculiventris abundance declined in more diverse landscapes, suggesting that it has more specialized habitat requirements. Egg mortality of H. halys by natural enemies was positively correlated with suburban habitats, while for P. maculiventris, it was correlated with soybean and grasslands. Lastly, stink bug egg mortality by natural enemies was highest adjacent to mixed annual crop plantings at a small, habitat-level scale.
The extension-focused project verified that H. halys populations on New Jersey orchards have declined over the past seven years and that growers and agricultural professionals no longer consider it the highest‑priority pest. Although H. halys remains a persistent economic concern, the decline in abundance coincides with the spread of T. japonicus and increasing natural enemy pressure.
Altogether, this project met its objectives and demonstrates that biological control of stink bugs is shaped by both habitat-scale and landscape-scale factors. For cooperating farms, these findings underscore the value of crop diversification. Importantly, the study confirmed that the adventive parasitoid Trissolcus japonicus is now established on all cooperating farms and that it parasitizes H. halys more than P. maculiventris and contributes meaningfully to biological control.
Education & outreach activities and participation summary
Participation summary:
Results from the pilot study were shared with research scientists by Emma Waltman at the Entomological Society of American Eastern Branch conference in March 2023. The talk was titled ‘Trophic interactions of native and invasive stink bug and their parasitoids depend on habitat context.’ Additionally, she shared these results and 2023 fieldwork plans at an Ecology & Evolution Graduate Program student seminar in April 2023.
Results from the comprehensive, landscape-scale study and the extension project were shared with research scientists by Emma Waltman at the Entomological Society of American Annual Meeting in Portland, Oregon in November 2025. The talk was titled ‘A quarter century of BMSB in NJ: Population trends, management, and biological control of a devastating-turned-manageable pest’ and was in a symposium about the pest status of the brown marmorated stink bug.
Furthermore, results from the landscape-scale study were shared by Emma Waltman at the Graduate Research in Evolutionary Biology and Ecology Conference in New Brunswick, NJ in February 2026. The talk was titled ‘Native and non-native egg parasitoid wasps provide similar biological control of the brown marmorated stink bug, an invasive agricultural, but through different means.’
We made a one-page flyer to share results from the landscape-scale study and the extension project with New Jersey tree fruit growers. The flyer can be found in section 5. This flyer will be distributed to growers at an extension meeting this upcoming spring/summer and will be posted on the Rutgers Plant & Pest Advisory website
Project Outcomes
One of the results of this study was that egg parasitism and predation of H. halys eggs on peach trees was higher when the peaches are across from diversified mixed annual crop blocks than when they are across from mowed grass, other tree fruit (apples/cherries), or forested habitats. These mixed blocks often contained brassicas, nightshades, cucurbits, mints, Apiales, cut flowers, and other annual flowering plants, and are common on mid-Atlantic fruit and vegetable farms. With this in mind, we can inform growers that planting such plots across from tree fruit can improve biological control of H. halys, which can reduce the amount of insecticide required to control the pest and improve sustainability and profit of the farming operation. This information has been shared with the participating growers where the survey took place via distribution of a one-page fact sheet.
Another interesting result is that Trissolcus japonicus, or the samurai wasp, was well-represented, as it was detected on all 15 of the farms where research was conducted. This likely informs why H. halys population numbers went down between 2018-2025 and why the pest is a lower priority to growers and agricultural professionals in the region compared to the early-to-mid 2010s. However, parasitism rates of H. halys by T. japonicus are still generally low at 8.42%, and H. halys remains a moderate and persistent pest. Still, Trissolcus japonicus has only been in the area for about a decade, and it is possible that parasitism rates will continue to improve as they have since 2017 (1.3% in peach) as the wasp naturalizes over time.
Some growers have expressed interest in releasing T. japonicus on their farms. While this has not been approved at this time, this study will inform this decision if it is approved in the future. The non-target effect of T. japonicus on P. maculiventris was low, at only 1.63%, which suggests that the wasp will not severely harm the beneficial predator’s populations.
This project was the first that I fully designed independently, as my earlier PhD work involved work on my advisor’s funded projects. I learned a great deal about project management through planning and executing an intensive 11-week field season on 15 grower-operated farms, mentoring and supervising through closely working with four under graduate field, GIS, and molecular assistants, and scientific collaborating, through my collaboration with Tara Gariepy’s lab with Agriculture and Agri-food Canada. Additionally, I gained technical skills through conducting geospatial analyses in ArcGISPro, statistical analyses in R Studio, and molecular analyses such as chelex-based DNA extraction and multi-plex PCR. I will take these skills forward as I continue in a career in entomological research in agricultural settings.
To develop this project even further, I received 4 additional small grants to fund this work. These grants included 3 small grants from my graduate program totaling $6,500 and a $1,000 travel grant from the Rutgers School of Graduate Studies to attend a conference and report my research. With the funding from my graduate program, I was able to visit Dr. Gariepy’s lab in Canada and process my samples.
From my communication with growers and from the time I spent at my farm sites, I learned about just how many decisions growers make that may impact their pest and beneficial insect populations. These include habitat management in non-crop surrounding areas and management decisions that I did not necessarily account for in this project, like spray frequency, mowing, and thinning. Seeing the variation of practices on farms underscores the importance of using large sample sizes when conducting research on operational farms. Additionally, I understand how what “works” on one farm to improve biological control may not work on another farm because of variation in surrounding landscape or due to different cultural management practices. I plan to continue researching similar questions as I did in this project, and hope to apply to many more SARE grants during my research career.