Report for GNE24-315
Project Information
This project investigates the potential of co-cultivating grapevines and a new variety of blight-tolerant transgenic American chestnuts in Northeastern United States agroforestry settings to provide a novel product and diversify the agricultural practices of this region’s vineyards and orchards. Given the ecological and economic significance of chestnuts and grapes, this research explores the viability of their joint cultivation within the Northeast by building upon traditional agroforestry practices observed in other geographic regions. The pairing of these two crops with distinct water use strategies is premised on the hypothesized niche facilitation of grape for chestnut: Grapes have large xylem vessels that uptake vast quantities of water, which may enhance the physiological performance of moderately drought-tolerant American chestnut under conditions of excess soil moisture. American chestnut physiology remains under-examined, given this species’s extirpation prior to the emergence of physiological ecology as a discipline. To address this hypothesis and knowledge gap, I propose research which aims to examine how conditions of competition versus facilitation, induced by co-cultivation under varying grape densities versus standard monoculture plantings, influence light saturated photosynthetic rates, leaf water potential, and stomatal conductance of each crop, as well as chestnut basal diameter and height and grape harvest yield. This research and a coinciding outreach strategy to local orchardists, vignerons, nut growers, and other farmers and agricultural collaborators, will occur over the 2024 and 2025 growing seasons, providing insight into the feasibility and benefits of co-cultivating chestnuts with grapes in the Northeast.
- Growth and Harvest Yield: To understand how grape planting density in this co-cultivation framework influences chestnut and grape growth performance compared to traditional monoculture systems, I conducted a growth assessment of grapevines and chestnut trees. I measured and assessed:
- Chestnut basal diameter
- Chestnut height
- Grapevine cluster counts per vine
- Grapevine cluster weights per vine
- Grapevine shoot lengths per vine
- Growth differences among treatments
- Photosynthesis and Water Relations: To determine the role of water usage as a driver of competition versus facilitation, I evaluated the photosynthetic capacity and water relations of grapes and transgenic American chestnuts. I measured and assessed:
- Leaf gas exchange, specifically light saturated photosynthetic (Anet) rates and stomatal conductance (gs)
- Leaf water potential
- Photosynthetic and water use differences among treatments
- Agricultural Outreach: To promote the adoption of this regionally novel co-cultivation method within Northeastern vineyards–e.g. the wine region of the Finger Lakes–I visited local farmers and presented my work to stakeholders and organizations within ESF’s existing network of outreach and communications. I led and participated in:
- Networking and professional engagement opportunities
- I led a series of tours of the grape and chestnut research plot for the Northern Nut Growers Association and Chestnut Growers of America 2024 Annual Joint Conference
- I led tours of and shared my research on the LRES grape and chestnut plot at the NE-2333 Conference at SUNY-ESF
- ESF site visits
- I led a tour of and shared my research on the LRES grape and chestnut co-cultivation plot during a site visit with the NY Chapter of the Master Teachers
- External site visits with two local orchards, Z's Nutty Ridge and Finger Lakes Nut Farm
- Networking and professional engagement opportunities
The purpose of this project is to investigate how co-cultivating grapevines and transgenic American chestnuts in an agroforestry setting can promote sustainable agricultural practices that enhance ecological resilience and economic viability in the Northeast region. Transgenic ‘Darling’ trees have been engineered with an oxalate oxidase (OxO) gene from wheat, thereby expressing an enzyme that confers a degree of tolerance to the fungal pathogen Cryphonectria parasitica (Newhouse et al., 2021). American chestnuts and grapes were selected as the focus organisms of this research, given the major ecological, social, and economic role chestnuts served prior to their decline from chestnut blight; the present-day high commodity value of grapes; and the fact that species of the genus Vitis are often lianas that naturally grow on trees (Keller, 2020). Moreover, there is an established agroforestry tradition of using hardwood trees as tutors for grapevines in Mediterranean and tropical climates (Altieri and Nicholls, 2002; Dupraz and Liagre 2019; Paris et al. 2019; Oliva Oller et al., 2022), though the viability of this system within a mesic environment, characteristic of our Central New York study site, has not been fully examined. The potential of mixed-species agroforestry to enhance growth and physiological performance of co-cultivated organisms and drive economic diversification in agricultural landscapes underscores the motivation for this research.
Co-cultivation of grapevines and American chestnuts may address several key issues in sustainable agriculture. First, growing grapevines on chestnut trees may yield savings on infrastructure. Chestnut trees require modest inputs, such that their maintenance may offset the costs of building and maintaining traditional grape trellis systems. Secondly, this project promotes the diversification of land use and income sources, potentially improving farm resilience to climate change driven market fluctuations. For instance, vignerons–i.e. farmers who cultivate a vineyard for winemaking–could receive supplementary income from the co-cultivated transgenic chestnut trees, which reliably mast if they can adequately tolerate chestnut blight. Further, this form of multilayer farming optimizes and increases the productivity of a plot, as the vertical scaffolding of the trees allows more crop to be grown on a given piece of land. Lastly, this agricultural framework can contribute to the restoration and conservation of the American chestnut, a species of substantial historical, ecological, and economic importance in the Northeast.
I propose to utilize a pre-existing unique experimental platform to test the relative benefits of co-cultivation relative to traditional monoculture in field conditions. Species interactions in co-cultivated settings can result in benefits to one or both species via facilitation or reductions in performance via competition. The degree of facilitation versus competition is likely to be density-dependent. The SUNY ESF Grape and Chestnut Co-Cultivation study, established in 2020, allows for testing of these interactions through a traditional agroforestry vineyard schema in which three plots, each containing nine trees trellised with varying densities of grapes, border a square crop field. Transgenic American chestnuts are planted alongside grapevines at densities of one, three, and five vines per tree, with nearby, non-paired chestnuts and grapes serving as monoculture controls (Figure 1 in Media). The physiological assessment of the chestnut trees will include LI-COR LI-6800 system measurements of light saturated photosynthetic rates, stomatal conductance, and leaf water potential across each grape planting density. A complementary growth assessment within this experiment will measure grape harvest yield and tree basal diameter and height to determine how the measured physiological processes may be influencing biomass acquisition and allocation, and whether co-cultivated crops may be facilitating or impeding the productivity of the other compared to monoculture plantings. We will not measure chestnut harvest yield, as the four-year-old chestnuts are unlikely to reach reproductive maturity by the conclusion of this study.
This project aligns with Northeast SARE's outcome statement by contributing to the goal of promoting sustainable agriculture practices that enhance environmental stewardship, farm profitability, and rural community vitality. This research honors the holistic connections among land, water, air, and all living beings through its consideration of plant-soil feedbacks, interspecies water relations, chestnut's pronounced carbon sequestration potential, and the ample socio-economic benefits that pairing chestnut trees with grapes may have for farmers and rural community members. By demonstrating the potential of mixed-species agroforestry to address complex agricultural challenges while also fostering ecological resilience and economic viability, our research will support the overall objectives of sustainable agriculture initiatives in the Northeast region. Moreover, by providing evidence of the benefits and challenges associated with this framework, our research will inform land managers and other agricultural stakeholders, and serve as a proof of concept toward promoting the adoption of these sustainable agroforestry practices.
Research



-
- Growth and Harvest Yield: This field site is located in Syracuse,
NY at the SUNY ESF Lafayette Road Experiment Station’s
Grape and Chestnut Co-Cultivar study, which was established
in 2020. The field work proposed here will take place
during the 2024 and 2025 growing seasons. This study’s
experimental design features three plots, each containing
nine four-year-old transgenic trees planted with densities
of 1, 3, or 5 grapevines around a square garden plot,
alongside grapevines and chestnuts grown individually
(Figure 1). Given this experimental design, I will analyze
data from this work with a split-plot analysis of variance
(ANOVA) with a main plot factor containing two levels: each
crop grown together or alone; and a sub-plot factor of
density of grapevines per chestnut (1, 3, and 5 vines). Our
response variables are remnant grape cluster count, remnant grape cluster weight, grapevine shoot length, and chestnut
basal diameter and height growth (Significant die back of the fruits on our vines, likely due to a mildew infection, necessitated an amended harvest yield methodology, which had formerly incorporated grape harvest yield as the weight of fruits and clusters per vine). This growth assessment will allow us to determine how physiological processes may
be influencing biomass acquisition and allocation under
conditions of competition versus facilitation.-
- Chestnut basal
diameter: At the end of the growing
season (October 2024), I measured the basal
diameter of each American chestnut tree (N = 40)
using a digital dial caliper down to the nearest
tenth of a millimeter. Each tree’s basal diameter
was measured twice at perpendicular angles,
then averaged to produce one basal diameter value
per tree.- I was forced to relocate out of New York State in August 2025, so this method was not able to be continued
during the 2025 growing season. I was not able to repeat this method and consequently calculate growth
rate across the 2024 to 2025 growing seasons, as I had originally planned and wished.
- I was forced to relocate out of New York State in August 2025, so this method was not able to be continued
- Chestnut height:
In October 2024, I measured chestnut stem
length for each American chestnut tree (N = 40)
with a tape measure as the distance from the stem
base to the tip of the apical bud after
straightening each tree by hand.- I was forced to relocate out of New York State in August 2025, so this method was not able to be continued
during the 2025 growing season. I was not able to repeat this method and consequently calculate growth
rate across the 2024 to 2025 growing seasons, as I had originally planned and wished.
- I was forced to relocate out of New York State in August 2025, so this method was not able to be continued
- Grape cluster count: In September 2024, I
counted the remnant grape clusters per vine for all grapevine planting densities. I trimmed off all clusters from the vines to reduce spread of fungal inoculum. - Grape cluster weights: In September 2024, I weighed all fruitless, remnant clusters per vine, following trimming, using a portable scale.
- Grapevine shoot lengths: In September 2024, I measured all shoot lengths per vine using a flexible measuring tape. Shoot lengths were measured as this season's vegetative growth, from the base to the apical tip of each shoot.
- Grape Harvest Yield: Given that I was forced to relocate out of New York State in August 2025, I was unable to use the Harvest Cluster Weight methodology outlined in Sabbatini et al. (2012), as originally planned. I was unable to apply this method to all fruit-producing grapevines and average values for sub-replicates (i.e. when 3 or 5 grapevines per chestnut), N ≥ 40.
- Growth and harvest yield differences among
treatments: Split-plot ANOVA with type
III sums of squares using lmer package of R with a
main-plot factor of cultivation method (mono vs.
co) and a sub-plot factor of grape planting density
(1, 3, or 5 grapevines per tree).
- Chestnut basal
-
- Growth and Harvest Yield: This field site is located in Syracuse,
- Photosynthesis and Water Relations: The physiological assessment of the
grapevines and chestnut trees included LI-COR LI-6800
system measurements of light saturated photosynthetic rates
(Asat) and stomatal conductance
(gs), and Scholander-type pressure chamber
measurements of leaf water potential across each grape planting
density level and monoculture planting. These measurements
took place across three periods of the growing season (early,
mid, and late summer) thereby affording comparisons of
physiological responses to conditions of excess soil moisture
and drainage. While predawn leaf water potential was initially proposed, we omitted it as the project progressed.While it is common in plant water relations research to measure both predawn and midday leaf water potential, the exceptionally wet summer that we had and the limited amount of leaves on these relatively small plants led us to determine that sampling additional leaves for predawn leaf water potential was unlikely to provide more useful information, while imposing a cost of increasing the disturbance to the plants. Our midday leaf water potential measurements tell us that these trees were all well-hydrated with no significant water limitations, so there was no need to invoke predawn measurements to infer whether there was soil moisture limitation of our leaf water potential measurements.-
- Leaf gas exchange
measurements: I used a standard open-flow
gas exchange system (LI-COR LI-6800 with 6
cm2 leaf chamber) and set
chamber conditions to 500 μmol s−1 flow rate, 1200 and 300 μmol
m−2 s−1 Photosynthetically Active Radiation
(PAR), 420 μmol mol−1 CO2, Relative Humidity (RH)
40–60%, and block temperature 20 °C. The two distinct PAR values mirror varying conditions of light and shade within each vine-tree co-cultivation system. I measured two
grape and two chestnut leaves within each grape planting
density level (1, 3, 5), one leaf per each shade condition (PAR value), for each of the three
experimental plots and two grapevine leaves and two
chestnut leaves from six vines and six trees within the monoculture plots (N = 15
grapevine and 15 chestnut trees; Table 1 in Media) three times
per growing season over the course of 2024 and 2025
(May, July, and September). I selected mature, fully
expanded attached leaves for measurements. I
performed leaf gas exchange measurements during the
mid-morning to early afternoon (9am -2 pm) to ensure
measurement parameters match environmental and
physiological conditions. - A subsample of leaves was sent to Brookside Laboratories to be measured for N content to assess
potential impacts of soil conditions on photosynthetic
capacity. Samples were intended to be dried and ground and analyzed
via combustion elemental analysis. However, Dr. John Drake and I received the surprising news that the lab was unable to complete analyses because there was not enough material in each sample packet. I incorporated material according to Dr. John Drake's specifications, and it is unclear to me if he had encountered a similar challenge with this lab. Given that I was forced to relocate out of New York State in August 2025, I was unable to collect and submit additional plant material. - Leaf water potential: I used a Scholander-type pressure chamber to measure
midday leaf water potential. I detached all leaves assessed for gas exchange, directly following the gas exchange measurements, and placed each individually in a Ziploc bag containing a moist paper towel, which was then placed in a dark cooler until leaf water potential measurements could be carried out approximately one to three hours later. I measured each detached leaf for the midday water potential measurements. - Photosynthetic and
water use differences among
treatments:
I followed the same split-plot ANOVA approach as
described above (Objective 1d)
- Leaf gas exchange
-
- Agricultural Outreach: I
will help to advance a broader adoption of this unique
co-cultivation method and reduce a local agricultural reliance
on monoculture systems in vineyards, orchards, and other
farming systems by communicating my findings with stakeholders
and organizations within ESF’s existing network of outreach and
communications.-
- External site
visits:
During the 2025 growing season, I traveled to local nut growers and orchards to learn about their
existing operations and how this experimental framework
might translate to new sites. In particular, my goal was to strengthen pre-existing collaborations within
ESF’s network. Elizabeth Mae Kehas-Dewaghe, for whom a
Letter of Commitment is attached, has shared
her contacts from pre-established research
relationships, given that these contacts demonstrated
an eagerness to maintain ongoing collaborative
partnerships. During these site visits, I
learned about and discussed current operations with
agricultural collaborators to gauge the feasibility of
implementing our co-cultivation schema within new
sites. I visited Jeff Zarnowski at Z's Nutty Ridge on May 26, 2025, and Marvin Russell at Finger Lakes Nut Farm on June 27, 2025.- Jeff shared that a key barrier to the broader adoption of this co-cultivation framework involves the current lack of a proof of concept. He stated, "No one wants to be the first to try this, farmers may want to see others succeed first"
- Marvin shared that a key barrier to the broader adoption of this co-cultivation framework involves the requirement that farmers manage two harvests at once, with grapes ripening in September and chestnut harvest occurring very soon after in October. He also expressed that grapevines could grow too tall in the chestnut's crown, inhibiting an efficient and effective harvest.
- Networking and
professional engagement: I shared my work at the
Northern Nut Grower’s Association and Chestnut Growers of America's Annual Joint Conference
in July 2024, which was hosted at ESF’s campus.
Additionally, I presented my work at the September 2024
NE-2333 Chestnut Conference "Biological Improvement of
Chestnut through Technologies that Address Management
of the Species and its Pathogens and Pests," which
also took place at SUNY ESF. I was unable to travel to
present my work at the NY-TACF chapter’s annual meeting
in 2025. - ESF site visits: During the 2024 field season, I
participated in field tours scheduled by ACRRP’s
Outreach Coordinator and Distribution Manager, Adriana
Del Grosso. Specifically, I led a tour in October 2024 for local teachers participating in the Master
Teacher Program. ESF’s ACRRP also invited members
of the Northern Nut Growers Association for a tour in
late July 2024, which I assisted in leading. During the 2025 field season, I was unable to invite new
collaborators to the Lafayette Road Experiment
Station’s Grape and Chestnut Co-Cultivar study to offer
educational tours and demonstrations.
- External site
-
Chestnut Growth Assessment
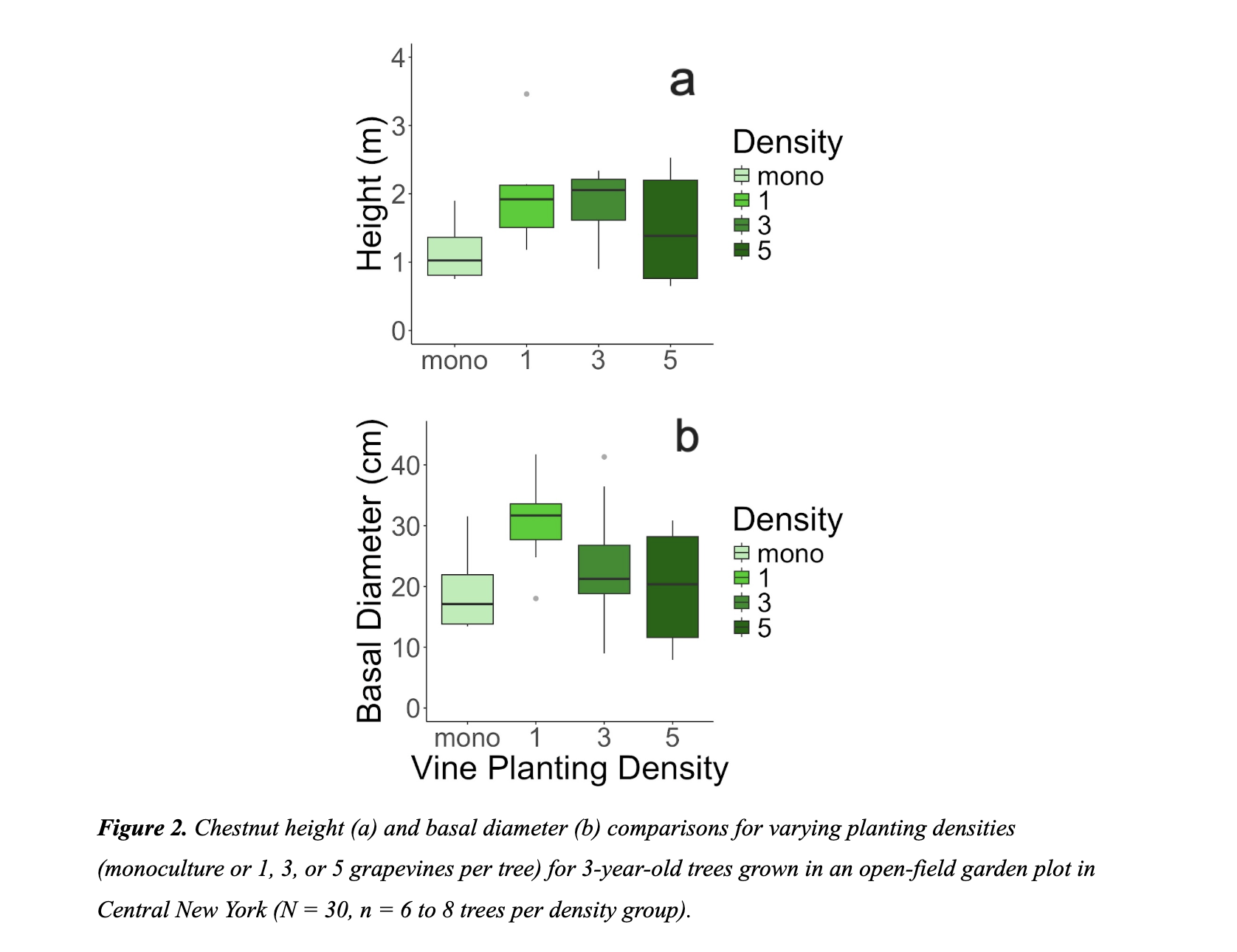
Observations of 3-year-old chestnut trees demonstrated that height did not vary significantly across the four grapevine planting densities (P = 0.12, ANOVA; Figure 2A), although we observed a modest increase in height for trees within the one and three vine planting densities. Chestnuts grown with a single grapevine were, on average, 80.9 cm (70.2%) taller than monoculture chestnuts, while trees grown with three vines were 68.5 cm (59.5%) taller. Likewise, there was no significant effect of density on basal diameter (P = 0.07; Figure 2B), although we observed a trend of a modest increase in basal diameter for trees planted in with a single grapevine (Figure 2b), which were 11.3 mm (58.6%) broader compared to monoculture trees. Ultimately, tree size was statistically similar across all density levels.
Grape Growth and Productivity Assessment
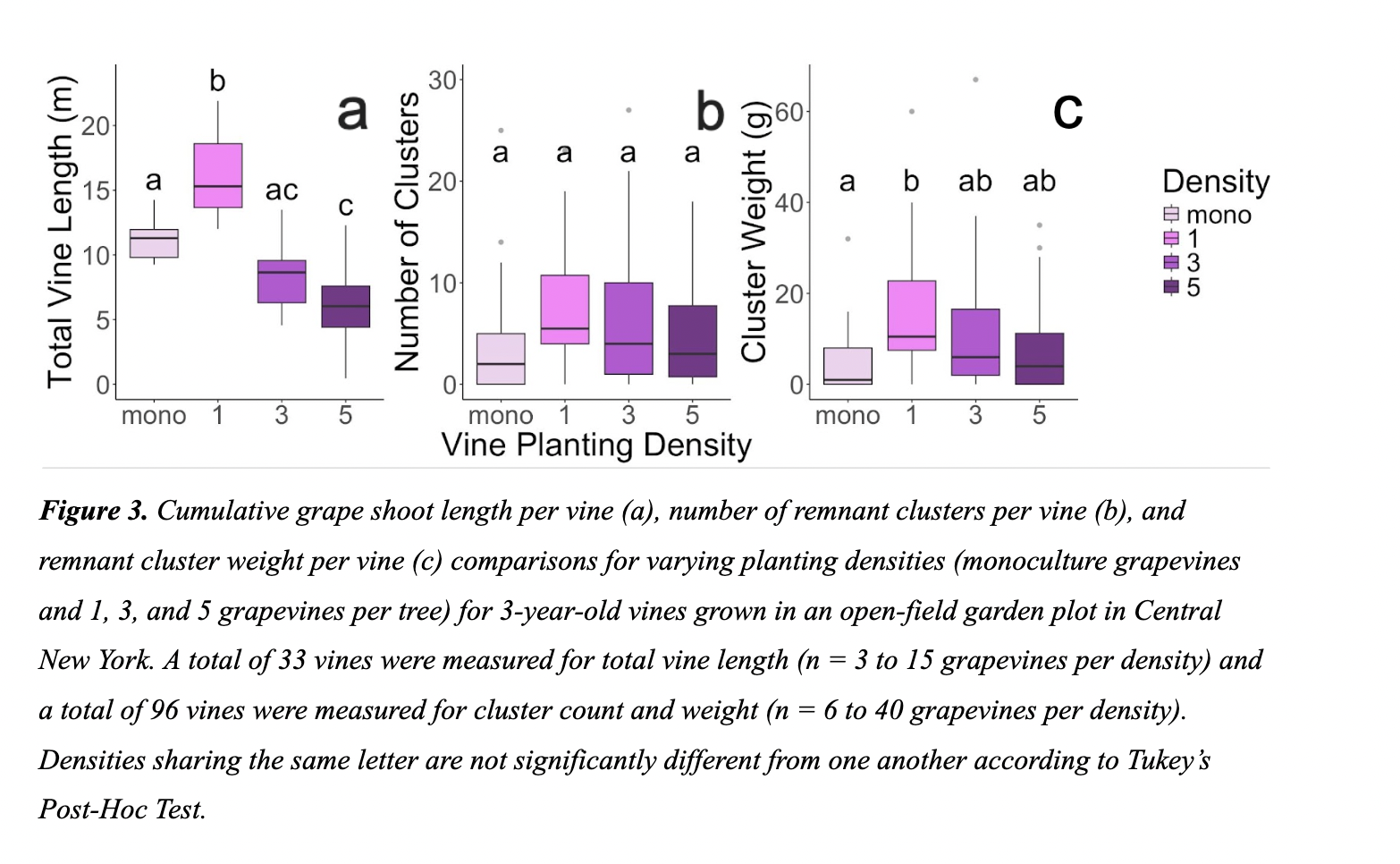
We observed a significant effect of density on total vine length (P < 0.001), such that single vines growing alongside a chestnut tree had, on average, 5.1 m (45.1%) greater cumulative vine length than monoculture grapevines (Figure 3a). Conversely, vines in the three and five density categories had reduced cumulative vine lengths than monoculture grapes (Figure 3a). In estimating productivity, we did not find a significant effect of density on the number of clusters per grapevine (P = 0.28), despite the modest trend of increased cluster count within the single vine density (Figure 3b), which produced, on average, 4.5 (109%) more clusters per vine than monoculture grape vines. As with total vine length, we found a significant effect of density on cluster weight per vine (P = 0.03; Figure 3c), where total cluster weight from vines in the single density group weighed 14g (280%) more, on average, than clusters from monoculture vines. Overall, vines in the one density grouping generally outperformed monoculture vines in terms of growth and productivity. We note that the data on chestnut growth (Figure 2) and vine growth (Figure 3) are both suggestive of a trend of enhanced performance in co-cultivation at low density, although this trend was not statistically significant in all instances.
Photosynthesis and Water Relations Assessment
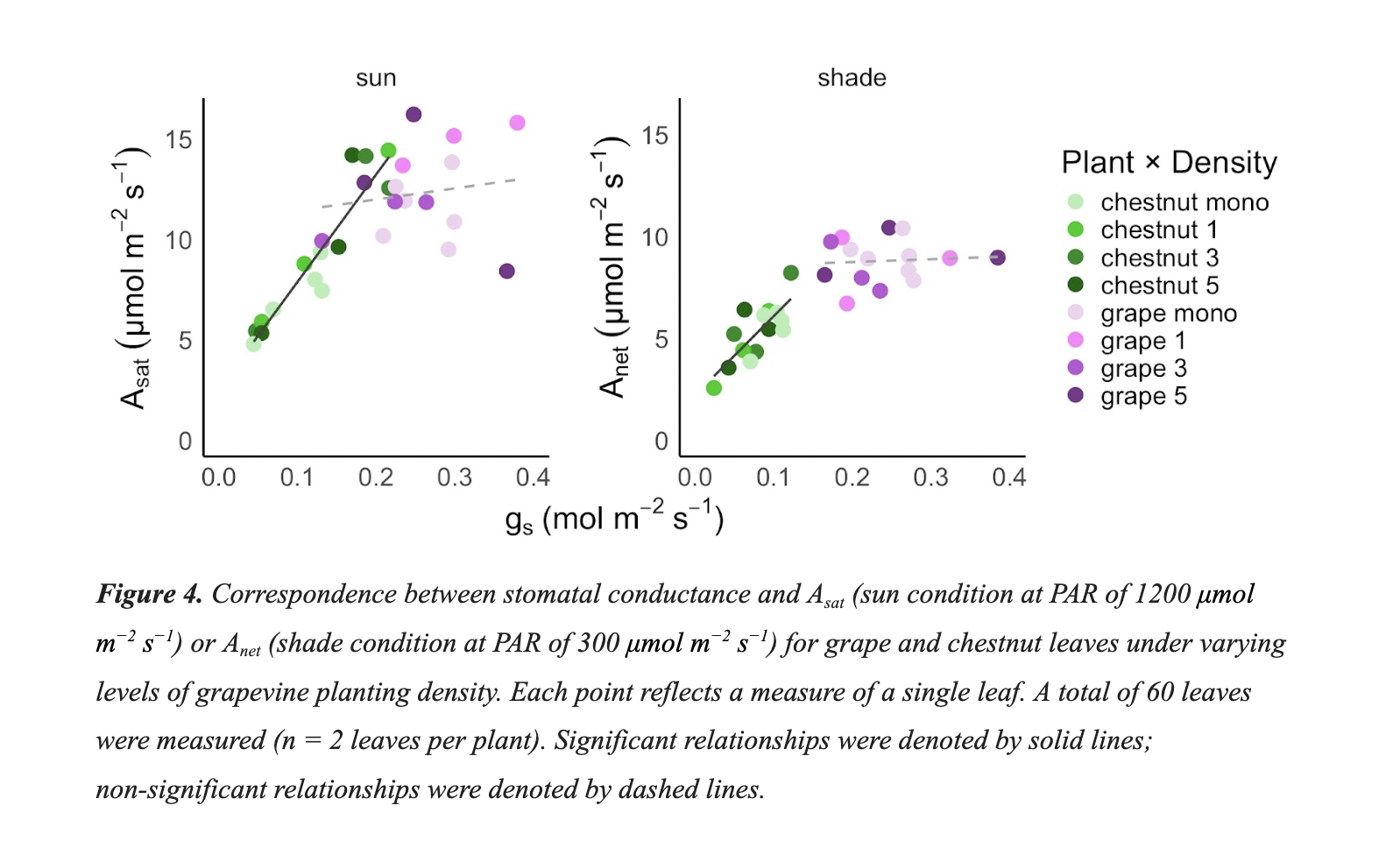
As expected, we found a significant main effect of plant type on photosynthetic rates reflective of the strong photosynthetic differences between American chestnuts and grape (Asat and Anet; P = 0.006 and P < 0.001, respectively). There was no significant main effect of density, (term not significant, Table 2). Grapevine Asat was 36% higher, on average, than Asat for chestnut and grapevine Anet was 66% greater than chestnut Anet (Table 3). We found no significant interaction between plant and density treatments, such that the magnitude of the differences in photosynthetic rates between each plant did not vary by density (interaction term not significant, Table 2).
Likewise, we found a significant main effect of plant type on gs for both the sun (P < 0.001) and shade (P < 0.001) treatments, such that gs varied significantly for each plant (Table 3). There was no significant main effect of density for either light condition, and no significant interaction between treatments (terms not significant, Table 2).
For ψleaf, there was no significant main effects of plant, density, nor treatment interactions (terms not significant, Table 2) for either light condition. Therefore, ψleaf did not vary significantly across plants and density levels (Table 3). Values of ψleaf were moderate across all observations, reflecting plants that had ample access to soil moisture with no discernable drought conditions.
Table 2. Linear mixed-effect model analysis of variance of plant, density (mono, 1, 3, and 5), and plant x density treatment effects on photosynthetic rates, stomatal conductance, and leaf water potential in Castanea dentata and Vitis vinifera under varying light conditions.
|
Plant |
Density |
Plant x Density |
|||||
|
F |
P |
F |
P |
F |
P |
||
Sun treatment |
Asat |
9.54 |
0.006** |
1.37 |
0.27 |
0.78 |
0.52 |
|
gs |
32.62 |
<0.001*** |
0.46 |
0.71 |
1.08 |
0.38 |
|
|
ψleaf |
0.63 |
0.44 |
0.81 |
0.50 |
1.46 |
0.25 |
|
Shade treatment |
Anet |
48.1 |
<0.001*** |
0.43 |
0.73 |
0.48 |
0.70 |
|
gs |
75.91 |
<0.001*** |
0.61 |
0.62 |
0.63 |
0.60 |
|
|
ψleaf |
0.10 |
0.76 |
0.26 |
0.85 |
0.06 |
0.98 |
|
Table 3. Photosynthetic rates, stomatal conductance, and leaf water potential (mean and standard error) in Castanea dentata and Vitis vinifera under varying light conditions. Densities sharing the same letter are not significantly different from one another according to Tukey’s Post-Hoc Test.
|
Plant |
Asat or Anet (μmol m-2s-1) |
gs (mol m-2s-1) |
ψleaf (MPA) |
|
Sun treatment |
chestnut |
9.03±0.93 a |
0.12±0.02 a |
-0.70±0.04 a |
|
grape |
12.31±0.60 b |
0.25±0.02 b |
-0.74±0.04 a |
|
Shade treatment |
chestnut |
5.29±0.39 a |
0.08±0.008 a |
-0.67±0.04 a |
|
grape |
8.81±0.28 b |
0.24±0.015 b |
-0.65±0.04 a |
We observed that photosynthetic rates were strongly dependent on gs for chestnut, but not grape (Figure 4). The regression slope line representing the relationship between photosynthetic rates and stomatal conductance was significant for chestnuts under both the sun (P < 0.001) and shade (P = 0.001) conditions, whereas the regression lines were not significant for grapes under either light treatment. We observed no difference in the nature of these regressions across densities (i.e., interaction terms not significant, Table 4).
We fit the Medlyn stomatal model (Medlyn et al., 2011) to this dataset and estimated the g1 parameters to be 3.0 for chestnut trees and 5.0 for grapes, the former of which is intermediate compared to other angiosperm trees in a global analysis (Lin et al. 2015). The 95% confidence intervals for these estimates did not overlap (2.7 to 3.3 for chestnut trees, 4.4 to 5.7 for grapes). Therefore, chestnuts exhibited greater WUE than grapes.
Measurements of gs were not dependent upon ψleaf for both grape and chestnut under both light conditions (Figure 5). However, chestnut plants under the shade condition exhibited a modest trend of decreasing gₛ with declining ψleaf, potentially indicative of more conservative stomatal regulation.
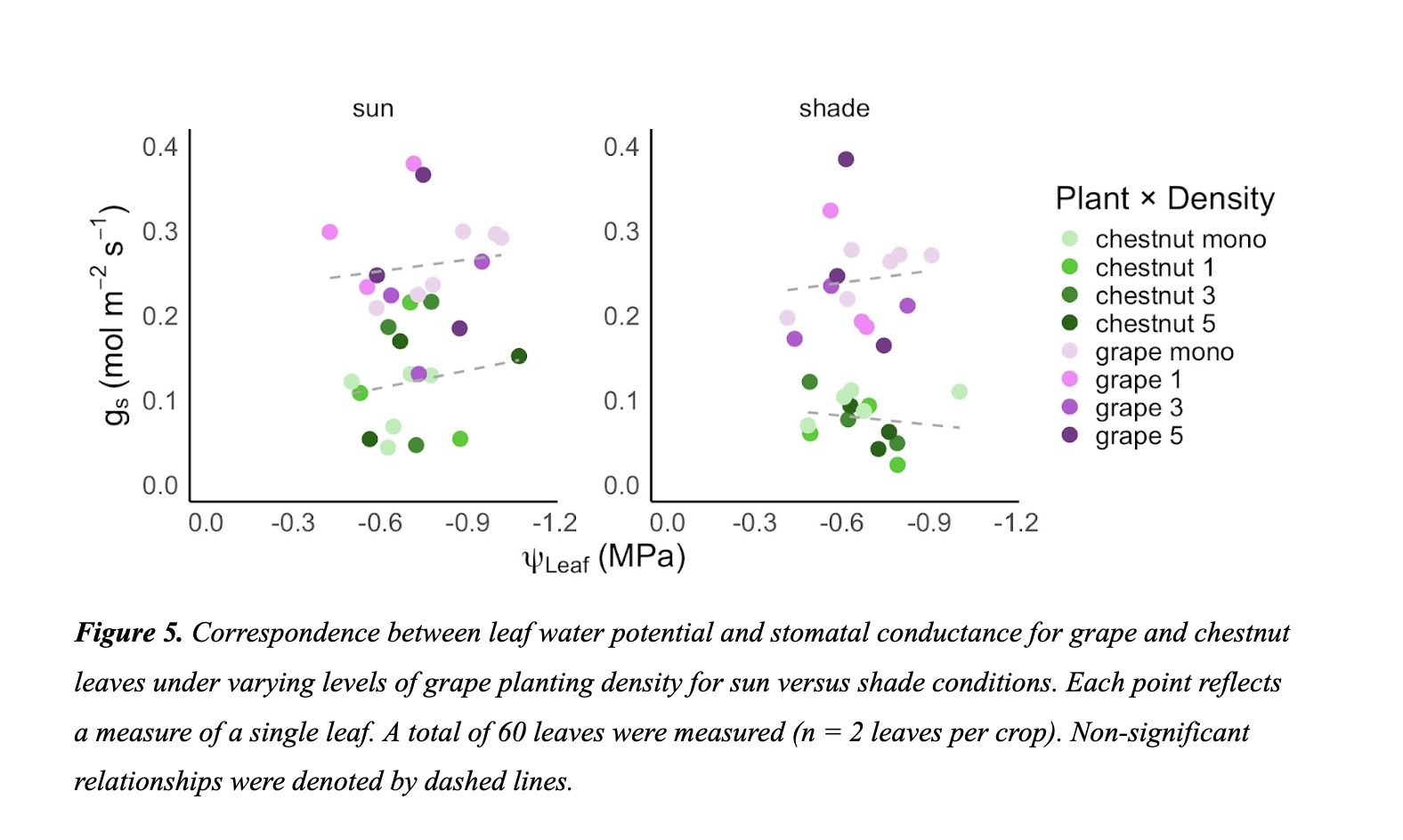
Table 4. Linear mixed-effect model analysis of covariance (ANCOVA) gs effects on photosynthetic rates (A) and ψleaf effects on gs in Castanea dentata and Vitis vinifera under full sun versus shade light conditions.
|
chestnut.sun |
chestnut.shade |
grape.sun |
grape.shade |
|||||
|
F |
P |
F |
P |
F |
P |
F |
P |
|
|
A ~ gs |
87.51 |
<0.001*** |
18.49 |
0.001** |
0.32 |
0.58 |
0.08 |
0.79 |
|
gs ~ ψleaf |
0.34 |
0.57 |
0.36 |
0.56 |
0.19 |
0.67 |
0.15 |
0.70 |
No significant differences in SLA were detected across density treatments for either chestnut or grape under sun or shade conditions, with the exception of grape shade leaves, where SLA increased as vine planting density increased (Table 5). Still, for chestnut sun and shade leaves, there was a modest trend of increased SLA between chestnuts planted with a single grapevine and the 3 and 5 vine density groups.
Table 5. SLA (cm2g-1) for chestnut and grape sun and shade leaves by density condition. Densities sharing the same letter are not significantly different from one another according to Tukey’s Post-Hoc Test.
|
SLA (cm2g-1) |
||||
|
Plant x Light Condition |
Mono |
1 |
3 |
5 |
|
chestnut.sun |
115.73 a |
106.81 a |
120.42 a |
126.60 a |
|
chestnut.shade |
156.23 a |
139.71 a |
168.48 a |
163.30 a |
|
grape.sun |
143.53 a |
149.92 a |
164.33 a |
145.95 a |
|
grape.shade |
150.67 a |
167.75 ab |
185.61 b |
171.51 b |
We assessed the growth of transgenic hybrid American chestnuts and growth and productivity of grapevines within a co-cultivation framework. Our results were indicative of asymmetric competition, in that co-cultivation strongly affected the vegetative growth of grapevines in a density-dependent manner, with more limited impact on American chestnut. Although mean chestnut height and mean basal diameter did not vary significantly across the varying vine planting densities, we found a non-significant increase in mean chestnut height for trees growing in the one and three vine planting densities when compared to monoculture chestnuts, and a modest increase in chestnut basal diameter for trees grown with a single vine. Thus, there may be a real impact of density on chestnut performance that this study could not detect because of our relatively modest statistical replication. Observed growth and productivity differences were not informed by our photosynthesis data, as we did not find a density effect on photosynthetic rates, gs, or ψleaf for either crop under either light condition. As expected, we observed a plant effect on photosynthetic rates and gs, such that grapevines performed higher rates of photosynthesis and exhibited greater stomatal conductance than chestnuts under both light conditions. Overall, these findings further our understanding of presently under-examined American chestnut physiology by characterizing it within the context of a well-studied crop–wine grapes. These results also suggest that co-cultivated grapes and chestnuts at the single vine planting density exhibit mutual, albeit asymmetric, facilitation, with a discernible shift toward competition at the three and five vine planting densities for grapes. Collectively, these results are supportive of the viability of their joint cultivation within the Northeast, as premised by traditional agroforestry practices observed in other geographic regions.
The asymmetric facilitation we observed may relate to the varying physiological responses of grape and chestnut in response to resource limitation. Although interspecies competition tends to increase under favorable environmental conditions, such as those present through the duration of this study, (Holmgren et al., 1997; Koffel et al., 2021), chestnut’s pronounced morphological and physiological plasticity may allow it to withstand some of this competition. Under low light conditions, many shade-tolerant plants invest more biomass in aboveground tissues (Holmgren et al., 1997), and this biomass partitioning strategy has been documented for American chestnut (Belair et al., 2018; Evans et al., 2023; Wang et al., 2006). For instance, we observed a modest, albeit statistically insignificant, trend of increased SLA for chestnuts planted with three or five grapevines compared to those planted with a single vine–a common adaptive response in eastern deciduous tree species–which can increase photosynthetic rates under light limitation (Givnish, 1988; Gottschalk, 1994). Likewise, the lack of a density effect on gas exchange rates for chestnut observed here is consistent with the intermediate level of shade tolerance that has previously been proposed for American chestnut, where light response curves demonstrate that photosynthetic capacity remains high for this species even under light limitation (Belair et al., 2018; Evans et al., 2023; Joesting et al., 2007; Joesting et al., 2009; Onwumelu et al., 2022). Accordingly, growth has been shown to remain strong in American chestnut at PPFD values as low as 360 mmol m-2s-1 (32% full sunlight) (Wang et al., 2006). As such, American chestnut is able to tolerate the shade produced by substantial competition, such as what we observed through co-cultivation with varying numbers of grapevines, without facing a growth penalty. Conversely, chestnut growth suffers under substantial water limitation when light is not limiting (Baurle et al., 2006; Brown et al., 2014). Our study did not incorporate or experience drought conditions, limiting our ability to assess how each crop would respond and interact under water limitation. Likewise, we were unable to observe performance under excess soil saturation at this upland study site. Our Central New York site received 113.6 cm of precipitation over the year of this study, which is comparable to the 30-year mean annual precipitation of 101.2 cm (National Oceanic and Atmospheric Administration; 1991–2024; Station USW00014771). Future studies could incorporate irrigation to assess each crop’s response to excess soil saturation.
Concerning grapevine growth and productivity, total vine length increased substantially for grapevines in the single vine density compared to monoculture and other planting densities. Because we observed no statistically significant impact of density on leaf water potential for grapevines, water appeared to not be limiting for grapevines, and differences in access to light or other resources may explain variations in growth across density treatments. Excessive vegetative growth is often a sign that water and nutrient resources are overly abundant, and can reduce vine fruitfulness, decrease fruit quality, and increase fruit disease (Wolf, 2008). As such, it is possible that the reduction in vegetative growth that we observed at the three- and five-vine planting densities is not necessarily negative in the context of producing higher quality fruits, and that mixed-species competition during favorable environmental conditions through co-cultivation could help to achieve a more balanced vine. Accordingly, there was no statistically significant difference between grape cluster count and mass at the three and five planting densities compared to the single-vine density and monocultures. Moreover, while we did not observe a quantitative tradeoff in grapevine vegetative and fruit growth and the single-vine density, we were unable to assess if the increased vegetative growth impacted fruit quality. Grape quality results from a complex balance of sugar and acid levels (Wolf, 2008), and various co-planted partner species can affect these components in different ways (Steiner et al., 2021). Similarly, grape varieties differ in how they respond to such partners (Postles et al., 2013), highlighting the importance of selecting co-planted crop species based on the specific requirements of each grape cultivar (Downey et al., 2006). Additionally, these measurements did not differentiate between primary and secondary shoots, the latter of which tends to exhibit reduced fruiting potential compared to primary shoots (Wolf, 2008).
This research also sought to characterize chestnut photosynthetic and water use physiology within the context of grapevine physiology, a well-studied crop. Grapevines are known to perform exceptionally high rates of photosynthesis with correspondingly high rates of water loss (Moutinho-Pereira et al., 2004). Transgenic American chestnuts performed relatively moderate rates of photosynthesis, even under the low light condition (Table 3), when also compared to documented maximum rates of photosynthesis described in Joesting et al. (2007) for seedlings of co-occurring forest tree species, such Acer saccharum and Betula populifolia, under various shelterwood canopy density treatments (Ellsworth and Reich, 1992; Wayne and Bazzaz, 1993), and when compared to other chestnut types (Knapp et al., 2014). Additionally, we reported increases in net photosynthesis with increased light, supporting previous findings that American chestnuts display plasticity in physiological responses to light (Evans et al., 2023; Joesting et al., 2009; Wang et al., 2006). Given the absence of a density effect on photosynthetic rates in either crop, the observed differences in growth may not reflect recent carbon acquisition but may instead reflect residual effects of physiological performance in previous growing seasons under unfavorable conditions.
Regarding American chestnut water relations, this work supports previous research by Bauerle et al. (2006) and Onwumelu et al. (2022) in characterizing American chestnuts as having relatively high WUE when compared to other tree species, and here, when compared to grapevines. Stomatal regulation of photosynthesis was stronger for chestnut than grape, in agreement with prior research into American chestnut gas exchange (Onwumelu et al., 2022). Further, grape plants showed little to no decline in gₛ as ψleaf became more negative, suggesting a relatively water-permissive strategy. In contrast, chestnut plants under shade exhibited a marginal, albeit statistically insignificant, decrease in gₛ with declining ψleaf, indicative of relatively more conservative stomatal regulation. However we caution that ψleaf values were all moderate, so we did not observe the full space of potential gₛ as ψleaf covariation. These trends suggest that grapevines may prioritize carbon gain under water-limited conditions, while chestnuts exhibit greater stomatal sensitivity to water status.
This study had several limitations. First, it was only modestly replicated with three co- cultivation plots established in one specific location in central New York. The facilitation or competition arising from co-cultivation is often context specific, depending on variables such as climate, topographical position, or soil conditions (Holmgren et al., 1997). A higher degree of replication would increase statistical power while also elucidating some of the context specificity of interspecific interactions. Secondly, we did not explicitly account for the actual light intensities experienced across the experiment, which has previously been found to strongly explain physiological responses to shading (Niinemets, 2007). Future studies should incorporate PAR measurements that capture the actual light conditions experienced by each co-cultivation unit during photosynthetic data collection. Thirdly, this study did not examine differences in growth performance across genetic backgrounds, which have been characterized for American chestnuts in previous work (Clark et al., 2012; Clark et al., 2023, Wegner et al., 2025). Moreover, we assessed height and basal diameter growth for chestnuts, although other aspects of morphology which are important within forest contexts differ within orchard settings. For instance, American chestnuts grown under substantial light limitation tend to exhibit reduced height growth and a broader crown morphology characterized by pronounced lateral branch growth (Wang et al., 2006). While the shorter morphology which may result from co-cultivation at high grapevine planting densities may be beneficial in orchard structural training where access to lower growing branches is prioritized, light limitation generally has a negative impact on nut yield and quality in chestnuts (Araki and Hall, 2000). Because this study was conducted early in the chestnut trees’ development, it is too soon to assess the effects of planting density on nut productivity. Additionally, it is too early to assess the blight tolerance of hybrid transgenic American chestnuts, or how infection may influence American chestnut growth. Long-term monitoring will be necessary to evaluate potential trade-offs between growth and blight tolerance across different co-cultivation densities.
Evaluating the viability of the enforcado vineyard agroforestry system in the northeastern U.S. will require continued analysis of interactions between wine grape and tree species under varying environmental conditions and competitive pressures. In this experiment, conditions were generally favorable, which may have intensified competition between crops, consistent with the stress-gradient response hypothesis. The relative competitiveness of grape alongside the stress tolerance of chestnut allowed each to persist under resource competition–most plausibly resulting from moderate light limitation. This pattern aligns with the complementary trait dynamics outlined in the refined Stress-Gradient Hypothesis (Maestre et al., 2009; Rolhauser et al., 2023). Still, it remains unclear whether grape and chestnut would facilitate each other’s survival under coinciding stressful conditions of light, water, and other resource limitations. Interestingly, prior studies have proposed that the enforcado system tends to experience a lower incidence of pest and disease attack compared to monoculture plantings, due to its enhanced habitat complexity (Oliva Oller et al., 2022; Oliva Oller et al., 2025). Ultimately, vegetative growth is not the sole indicator of overall plant performance in orchard and vineyard contexts, and future analyses of nut and fruit yield and quality will be important for advancing this framework in the Northeast. The joint cultivation of wine grapes and chestnuts not only has the potential to enhance each crop’s performance under increasingly variable climate conditions, but also to advance local biodiversity through tree species restoration. In contrast to forest restoration contexts, agroforestry systems are relatively controlled and managed, such that blight tolerant chestnuts can grow with reduced pressure from pests, pathogens, and competition. These controlled environments may allow for more precise selection of individuals that exhibit both vigorous growth and high nut quality during breeding efforts.
Education & outreach activities and participation summary
Participation summary:
This proposal’s outreach plan is outlined within the “Agricultural Outreach” Objective and Methods sections. In summary, this outreach strategy centers on conducting site visits to orchards, vineyards, and other relevant agricultural sites; presenting at annual conferences; and hosting site visits at the co-cultivar study to offer educational demonstration tours. My 2024 outreach strategy focused primarily on leading ESF site tours with organizations within ACRRP’s existing network of outreach and presenting at conferences. For instance, I led tours and shared my work in late July at the Northern Nut Growers Association and American Chestnut Growers Annual Joint Conference, in September 2024 at the NE-2333 Chestnut Conference "Biological Improvement of Chestnut through Technologies that Address Management of the Species and its Pathogens and Pests," and in October 2024 for local teachers participating in the Master Teacher Program. My 2025 strategy will focus on traveling to collaborators to learn about their existing operations and how this experimental framework might translate to new sites, but will also involve hosting site demonstration tours and presenting my research findings at more conferences and annual meetings. Unfortunately, I was unable to travel to present at the NY-TACF chapter’s annual meeting in Laurens, NY in 2025.
The goals of this outreach strategy are to strengthen pre-existing partnerships, learn more about local agricultural operations, and share the findings and lessons learned from my research. Ultimately, I would like to promote the broader adoption of this unique co-cultivation method within the Northeast, given that grape and chestnut co-cultivation has immense potential to address pressing challenges related to climate change and sustainable agriculture.
Project Outcomes
This project investigated the potential of co-cultivating grapevines and a new variety of blight-tolerant transgenic American chestnuts in northeastern United States agroforestry settings to provide a novel product and diversify the agricultural practices of this region’s vineyards and orchards. Given the ecological and economic significance of chestnuts and grapes, this research has the potential to advance awareness of the viability of their joint cultivation within the Northeast by building upon traditional agroforestry practices observed in other geographic regions. This research also helps to build upon our presently limited understanding of American chestnut physiology, given this species’ extirpation prior to the emergence of physiological ecology as a discipline, through its analysis of photosynthetic rates, leaf water potential, and stomatal conductance of each crop, as well as chestnut basal diameter and height and grapevine growth. By integrating ecological theory with sustainable agricultural practices, these findings aim to support chestnut restoration while enhancing the sustainability and economic diversification of Northeastern agroforestry systems. The judicious selection of sites for reintroduction which support American chestnut’s adaptive responses to stress, with a critical examination of how microsite variations influence chestnut physiological plasticity, thereby variably promoting interspecies competition or facilitation, will be critical for ensuring the success of blight tolerant chestnut during restoration efforts.
Overall, this work's motivation lies in advancing a sustainable co-cultivation framework which may help crops to be more resilient in the face of increasingly harsh environment conditions. Our hope is that co-cultivating grapevines and hardwood trees (such as chestnuts) may minimize irrigation needs; enhance photosynthetic capabilities--thereby potentially enhancing harvest yield--of each crop; and offer a novel agricultural product through the re-introduction of the presently functionally extinct American chestnut tree.
My awareness of sustainable agriculture has certainly expanded due to this project. I have learned a lot about the benefits of mixed crop co-cultivation and how compatible crops can improve each other's resilience during unfavorable weather conditions. I've honed my skills in measuring and assessing plant health and performance, and I worked hard to make sense of the data I collected. I've also had informative conversations with farmers and tree growers about potential challenges associated with scaling up the co-cultivation framework I have been studying, despite its potential benefits for crop health and productivity. Alongside these successes and meaningful conversations, I've also learned a lot about the every day challenges associated with growing crops, such as the threat of fungal infections, damaging effects of excess precipitation, and physical toll of long days spent out in the field collecting data. Still, this research opportunity has offered me connections in the industry I may not otherwise have had access to, which I am grateful for.