Progress report for GNE24-330
Project Information
Ants are ubiquitous in terrestrial ecosystems and play crucial roles in maintaining ecosystem functioning. In many agricultural systems, ants can provide valuable services, including pest control. In northern temperate crop fields, however, the presence and potential impact of ants has been largely overlooked, primarily due to widespread use of tillage. Increasing adoption of no-till farming, particularly in the mid-Atlantic region of the United States, presents an opportunity to study and harness the potential of ants for sustainable pest management. This project in Pennsylvania aims to examine the potential for another conservation practice, cover crops, to grow ant communities and their subsequent effects on pest control services in no-till corn and soy production. I will evaluate the effects of cover crops grown in monocultures and mixtures on abundance, species richness, and functional roles of ant communities in no-till fields. Furthermore, I will compare the effects of cover crop monocultures and mixtures on diversity of ant communities and the larger epigeal predator communities on the control of crop pests and crop damage. This research will contribute to development of sustainable pest management strategies that leverage ecosystem services provided by ants in no-till agroecosystems. The findings will have significant implications for sustainable agriculture, particularly no-till and cover crop use. The results from this study will help farmers reduce their resilience on synthetic insecticide by integrating cover cropping and integrated pest management (IPM) practices with no-till farming. This integration will enhance the potential for ants, and other predators to provide effective pest control services.
- Evaluate the effects of cover crop (monocultures and mixtures) on abundance, species richness, and functional roles of ant communities and species in no-till fields.
- Compare the effects of cover crop (monocultures and mixtures) on diversity of ant populations and the larger epigeal predator community across growing seasons.
- Under different cover crops (monocultures and mixtures), assess influences of changes in ant species diversity and abundance, and that of the larger epigeal predator community, on control of crop pests and crop damage.
The purpose of this project is to examine in no-till agroecosystems the impact of cover crops on ants and the larger soil-borne (i.e., epigeal) community of predatory arthropods and their effects on pest control services. Ants are ubiquitous, and their activities can define plant and animal communities [1,2]. In tropical and subtropical agriculture, ants are considered important natural enemies of arthropod pests as they protect crop plants against a wide range of herbivores. Their predatory behavior is facilitated by their aggressiveness and sophisticated social organization, which enables them to efficiently locate, capture, and transport the prey back to their nests. However, little is known for the Northern US Corn Belt about how they contribute to pest suppression, especially when combined with conservation practices like cover crops and integrated pest management. The functional roles of ants have largely been overlooked because historic use of tillage has limited presence and diversity of ants in large-acreage crop fields. By disrupting soil where they nest, tillage excludes ants from playing a significant role in the agroecosystems. In contrast, no-till farming enables ants to colonize fields and contribute significantly to pest management. Given evidence of the importance of ants in other agroecosystems, we expect them to be key contributors to pest control in no-till fields.
No-till farming practices, which minimize soil disturbance, can create favorable environments for ants to establish colonies in crop fields [2]. In regions like the mid-Atlantic US, no-till farming has been widely adopted over thousands of acres of field crops like corn and soybeans. This adoption has been largely driven by regional initiatives aimed at reducing sediment and nutrient runoff into the Chesapeake Bay. This shift, along with the adoption of conservation-based practices like cover cropping and integrated pest management (IPM), strengthens the opportunity for ants to become long-term residents and exert their influence on agroecosystems.
While no-till allows ants to colonize fields, the influence of cover crops on ant communities and their potential for pest control is unknown in the Northern US Corn Belt. Studies conducted in other agroecosystems suggest that cover-crop diversity may influence ant communities by providing improved habitat quality, offering a greater variety of resources such as food (e.g., seeds and insect prey; [3,4]), shelter (e.g., different plant architecture and litter layers,), and nesting sites (e.g., improved soil quality, structures, and microhabitat; [5]). These findings from other agroecosystems are suggestive of how cover crops may be influencing ant communities in the no-till agroecosystems of the Northern US Corn Belt. However, the specific details still need to be investigated as we have little information on the role that ants play in any form of current field-crop production across the Corn Belt. Increased habitat heterogeneity associated with cover crop mixtures may even support a higher diversity of ant species and functional groups than monocultures, potentially leading to more effective and resilient pest control services.
Pest damage is a major challenge in agroecosystems, causing an estimated ~20% loss in global crop production, amounting to ~$470 billion United States (US) dollars in annual loss worldwide [6]. Each year in the US, the financial impact of reduced crop yield by insect pests and expenses related to insecticide use surpasses $15 billion [7]. Traditional pest management strategies primarily depend on synthetic insecticides [8], which negatively affect the diversity of non-target arthropods [9,10], and present a concern for human and environmental health [11]. To safeguard future food supplies and maintain the stability of agricultural ecosystems, it is essential to create and implement eco-friendly pest control strategies that can serve as viable substitutes for pesticide-based approaches.
This project's findings will have significant implications for sustainable agriculture, particularly regarding conservation practices, including no-till, cover crops, and IPM. By identifying cover crop strategies that optimize ant-mediated pest control in no-till fields, this research will contribute to developing more effective and ecologically based pest management approaches tailored to these systems. However, it is crucial to recognize that ants will not be effective in no-till fields with cover crops unless farmers combine these practices with IPM strategies. The integration of no-till farming, cover cropping, and IPM will create a synergistic approach that enhances the potential for ants to provide effective pest control services. Implementing these strategies will help farmers reduce reliance on synthetic insecticides, which can cost up to $4.2 billion annually in the United States alone [7]. Enhancing these ecosystem services can lead to long-term economic benefits for farmers through increased crop yields, reduced input costs, and improved soil fertility [12].
Research
Field Set-Up (all objectives)
To achieve the three objectives, I will conduct a two-year field experiment at the Penn State’s Russel E. Larson Agricultural Research Center (Rocksprings, PA). To reduce legacy effects, I established the experiment in a field that has not been tilled for the last ten years. I established a randomized complete block design with five replicates of six cover crop treatments. To explore the influence of different cover crop species, I planted them as single species (i.e., monocultures) or mixtures of species. Monocultures of cover crop species were based on functional traits and include: red clover (Trifolium pratense L.), forage radish (Raphanus sativus L. ‘Tillage Radish’), and cereal rye (Secale cereale L. ‘Aroostook’). These cover crop species are known to be winter hardy (clover and rye) and susceptible to winter kill (radish) in central Pennsylvania. Mixtures of these species can contribute to weed suppression or nitrogen management [13]. Importantly, these cover crop species are used by farmers in Central Pennsylvania [14]. To explore the effects of each species alone and in mixtures, I planted monocultures of each of the three species and randomly assigned the three possible two-species mixtures across the experimental blocks (cereal rye + red clover; cereal rye + radish; red clover + radish), along with the three-species mixture. For a control, I included plots with no cover crop.
Each experimental plot was 15 x 15 m, arranged in a grid pattern with 5-m buffer zones between plots to reduce movement of arthropods among plots. Plots were grouped into five blocks, and each treatment was randomly assigned to one plot within each block, resulting in 30 plots (6 cover crop treatments x 5 replicates). The experimental plots will maintain consistent identities and sizes throughout the two-year experiment, allowing cumulative effects of treatments to emerge and be measured over the duration of the study. Penn State farm staff implemented the treatments using standard no-tillage farming practices and best management practices. Compared to cover-crop monocultures and the no cover-crop treatment, I anticipate that cover crop mixtures will enhance abundance, diversity, and functional roles of ants. I expect monocultures to support higher abundance and diversity than no cover crop plots. Further, I also expect that two-way and three-way mixtures will have a more pronounced effect, with the three-way mixture having the greatest impact due to increased habitat complexity and resource diversity.
In October of the first year (2024), I planted the different cover crop treatments. In the following spring, I evaluated biomass production of the treatments, then sprayed glyphosate in all plots to terminate the cover crop and kill weeds. Shortly after cover-crop termination, the plots was planted to maize. I used a non-Bt, glyphosate-tolerant hybrid (104 d maturity) that performed well in Penn State Extension’s Corn Variety Trials. After grain harvest in November, plots will be planted with the same cover crop treatments with biomass evaluated again in spring prior to termination. In the spring of the second year of the experiment, the field will be planted to glyphosate-tolerant soybean (maturity 3.0).
Objective 1. Evaluate effects of cover crop mixtures on abundance, species richness, and functional roles of ant communities and species in no-till fields.
Ant sampling. I used pitfall traps and ant baits in all the experimental plots to assess the epigeal arthropod community, including ants. To sample the epigeal community, in each plot, I placed two 16-oz deli containers flushed with the soil surface. Each container held a 50:50 mixture of water and propylene glycol to kill and preserve captured insects and spiders. Each month (July-September), I opened traps for 72 h. In the laboratory, I identified captured arthropods to the lowest possible taxonomic level. The pitfall trapping revealed when species were active and the influence of cover-crop treatments on their activity.
In addition to pitfall trapping for the whole epigeal community, I also used baits to further evaluate ant communities and activity. After each trapping session, I deployed two baiting stations in each plot, offering preweighed food items, including sugar water-soaked cotton balls, Pecan Sandies (these cookies are particularly attractive to ants), and dead waxworms; this diversity of baits attracted a wide range of ant species. I deployed baits at sunset and checked them after three and twenty-four hours. I collected unknown ant species to determine their identity in the laboratory.
Evaluating ant functional roles. To assess functional roles of ants in a no-till system, I deployed seed cards and conducted a sentinel prey experiment using waxworms (Galleria mellonella) to measure predation rates. I conducted this assay three times (July, August, and September), shortly after pitfall trapping. In each plot, I deployed 8 live caterpillars, with four placed inside each of the two cages (hardware cloth cylinder tops with a plastic lid) that excluded vertebrates (e.g., mice, birds). I pinned each caterpillar to the soil and checked on them after three and twenty-four hours to record prey removed and identify any predators that are present.
To measure the influence of ants on weed seeds, the morning after the caterpillar assay, I deployed in each plot inside the same exclusion cages one seed tray, each holding 40 seeds. Each card will contain 20 seeds of red rot pigweed (Amaranthus retroflexus) and yellow foxtail (Setaria pumila), affixed to a double-sided tape on a Petri dish and sprinkled with sand to prevent invertebrates from getting stuck. These weed species are common in central Pennsylvania and are known to be collected by ants and other seed predators (e.g., ants, carabid beetles, and crickets). I left these seed trays in place for 48 h and then counted the number of remaining seeds on each card.
Objective 2. Compare the effects of cover crop mixtures and single species cover crops on the diversity of ant populations and the larger epigeal predator community across different growing seasons.
To compare effects of cover crop treatments on ant diversity across over time, I used data collected via pitfall traps on ant diversity and the broader predator community. I calculated diversity indices, such as the Shannon index and Simpson index, for each treatment, month, and year. These indices provided a quantitative measure of the diversity of ant species and the broader predator community and accounted for both species richness and evenness. Furthermore, I employed appropriate statistical methods, such as Generalized Linear Mixed Models. These analyses helped determine if there are significant differences in ant diversity among single-species cover crops and cover-crop mixtures. The repeated measures design allowed for assessing changes in ant diversity over time and across different growing seasons.
Objective 3. Assess influences of changes in ant species diversity and abundance, and that of the larger epigeal predator community, with different cover-crop treatments on control of crop pests and crop damage.
Every other week on four randomly selected 3-m sections of row per plot, I quantified the damage that pests have caused to crop plants using a rating scale (0: no damage, 1: 1-25% damage; 2: 25-50% damage; 3: 50-75% damage; and 4: 75-100% damage; [15]).
Given the significant challenge slugs pose to Pennsylvania no-till farmers and the possibility that ants influence their populations, I will measure slug populations in each plot every other week using 0.1-m2 shelter traps made of white roofing shingles. Previous research from the Tooker lab has shown that slugs under shingles correlate with the extent of damage to plants. Slug damage severity will be assessed when they are most relevant (V2 and V5 growth stages) for maize and soybeans using a 0-4 rating scale 0: no damage, 1: 1-25% damage, 2: 25-50% damage; 3: 50-75% damage; and 4: 75-100% damage; [15]).
Statistical analysis
To assess the impact of cover crop treatments on ant communities, pest populations, and pest control services, I will employed generalized linear mixed effect models (GLMM). These statistical methods evaluated differences in ant abundance, species richness, functional roles, and the abundance and diversity of the epigeal predator community across treatments. To investigate the influence of changes in ant communities, and the larger predator community, on pest control, I will conduct regression analyses to explore the relationships between continuous variables such as ant abundance, diversity, predation rates, pest populations, and crop damage.
To ascertain the strength of associations between specific ant species and cover crop treatments, I will conduct indicator species analysis to identify groups of species or functional traits that may be related to a particular treatment. This analysis will provide insights into which ant species or functional groups are most responsive to cover crop mixtures and may contribute significantly to pest control services.
Project Overview
This project is evaluating how cover crop species and mixtures influence ant communities and their ecosystem services in no-till corn and soybean systems. We aim to identify cover crop strategies that enhance beneficial ant populations and their contributions to pest control and weed management in mid-Atlantic agroecosystems.
Project Objectives:
Objective 1: Evaluate the effects of cover crop monocultures and mixtures on ant abundance and their functional roles (predation, weed seed removal) in no-till agriculture.
Objective 2: Compare the effects of cover crop monocultures and mixtures on diversity of ant populations and the larger epigeal predator community across growing seasons.
Objective 3: Under different cover crops (monocultures and mixtures), assess influences of changes in ant species diversity and abundance, and that of the larger epigeal predator community, on control of crop pests and crop damage.
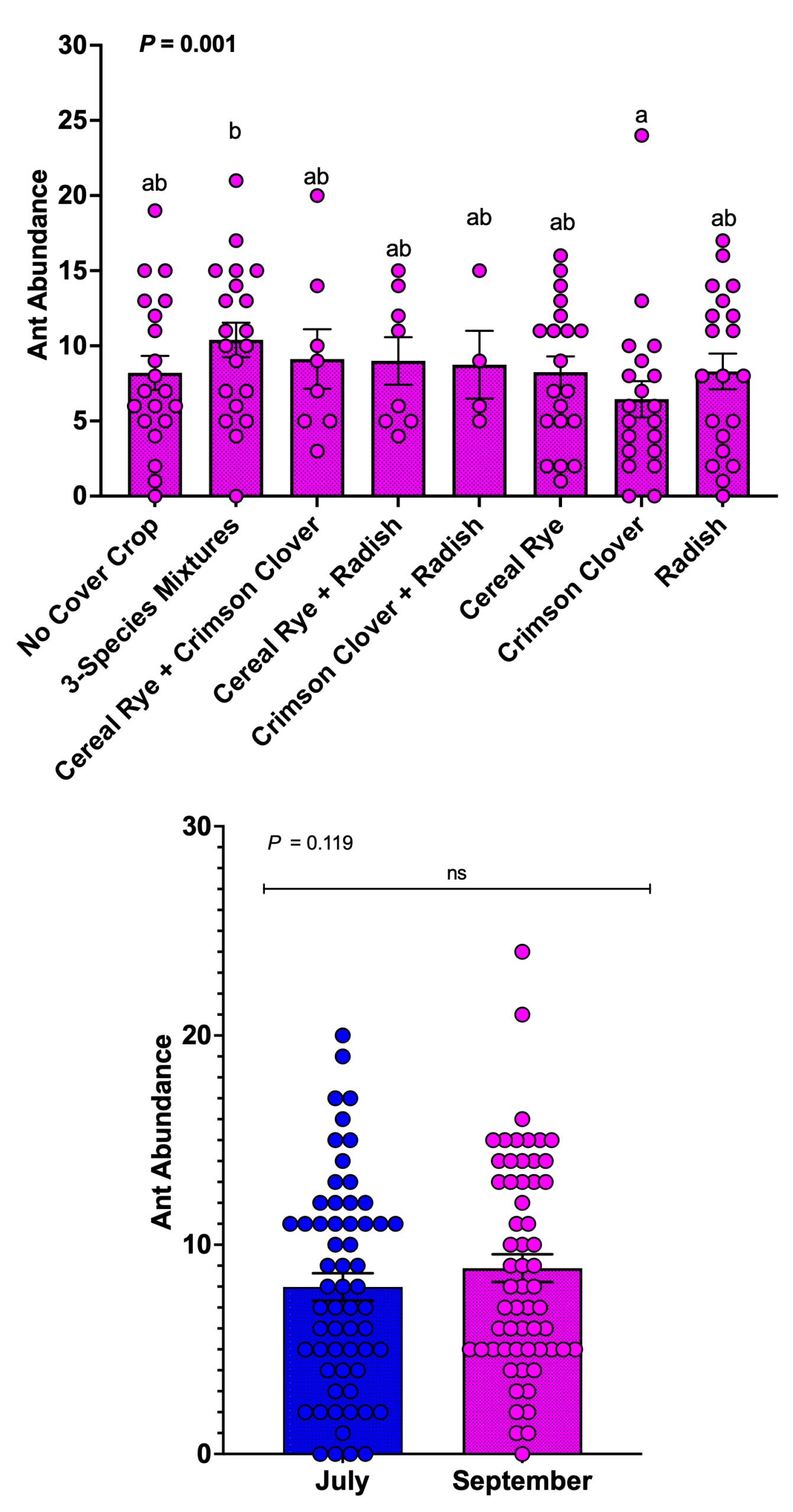
Epigeal arthropods and ant abundance in pitfall traps
We captured 1,377 epigeal arthropods in pitfall traps during the 2025 growing season. Ants dominated the epigeal predator community and comprised 73.5% of all arthropods captured (1,012 of 1,377). Other predators included ground beetles, spiders, tiger beetles, and harvestmen, which together accounted for 365 individuals.
Cover crop treatment significantly affected ant abundance in pitfall traps (P = 0.001). The three-species mixture supported the highest ant abundance and differed significantly from crimson clover alone. Most other cover crop treatments supported intermediate ant abundance and did not differ from one another or from the no-cover control. Contrary to our hypothesis, the no-cover control did not support the lowest ant abundance; instead, crimson clover alone supported fewer ants than several other treatments. These results indicate that cover crop identity, rather than the presence of cover crops alone, influences ant populations. Month did not affect ant abundance (P = 0.119), and ant activity in pitfall traps remained stable between July and September.
Ant foraging activities
Cover crop treatment, month, and bait type all significantly affected ant foraging activity at baits (Treatment:P < 0.001; Month: P < 0.001; Bait type: P < 0.001). The three-species mixture supported the highest ant foraging activity and differed significantly from the no-cover control, crimson clover + radish, and cereal rye + radish. Most other treatments supported intermediate ant activity and did not differ significantly from one another. These patterns may suggest that cover crop identity, rather than cover crop presence alone, shapes ant foraging responses.
Bait type strongly influenced ant visitation. Pecan Sandies attracted more ants than either waxworms or sugar water on cotton balls, which may suggest that ants in this system preferentially forage on carbohydrate-rich resources during the growing season, or that the cookie baits' combination of sugars and lipids was particularly attractive.
Ant foraging activity peaked in August and differed significantly from both July and September, with the lowest activity in July. This temporal pattern likely reflects seasonal changes in colony size and foraging intensity, with peak activity occurring in mid-summer.
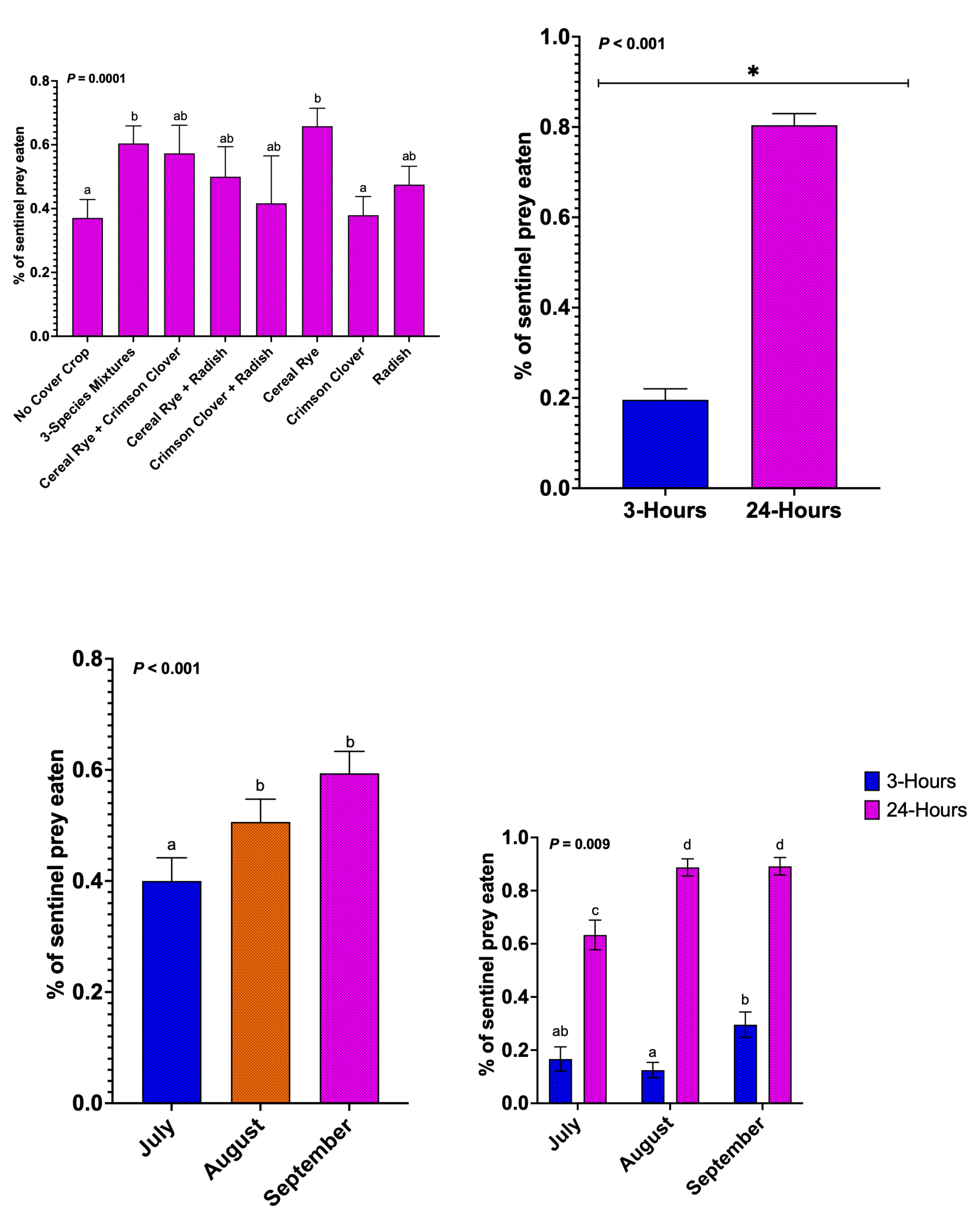
Predation of sentinel prey
Cover crop treatment, month, and time period significantly affected predation of sentinel waxworm (Beta-binomial GLMM; Treatment: P = 0.0001; Time: P < 0.001; Month: P < 0.001; Month × Time interaction: P = 0.009). Cereal rye and the three-species mixture supported the highest predation rates and differed significantly from no-cover crop control and crimson clover alone. Most other cover crop treatments showed intermediate predation rates.
Time period strongly influenced predation success. Waxworms exposed for 24 hours experienced significantly higher predation compared to those exposed for three hours. This pattern occurred across all months, although the magnitude varied seasonally (Month × Time interaction, P = 0.009).
Predation rates increased across the growing season for both time periods. July showed the lowest predation rates, followed by progressive increase through August and September. This seasonal increase likely reflects growth in ant colony and greater foraging activity as season progressed. Consistently higher predation during 24-hour time period suggests that longer prey exposure allows more time for predators to locate and consume prey.
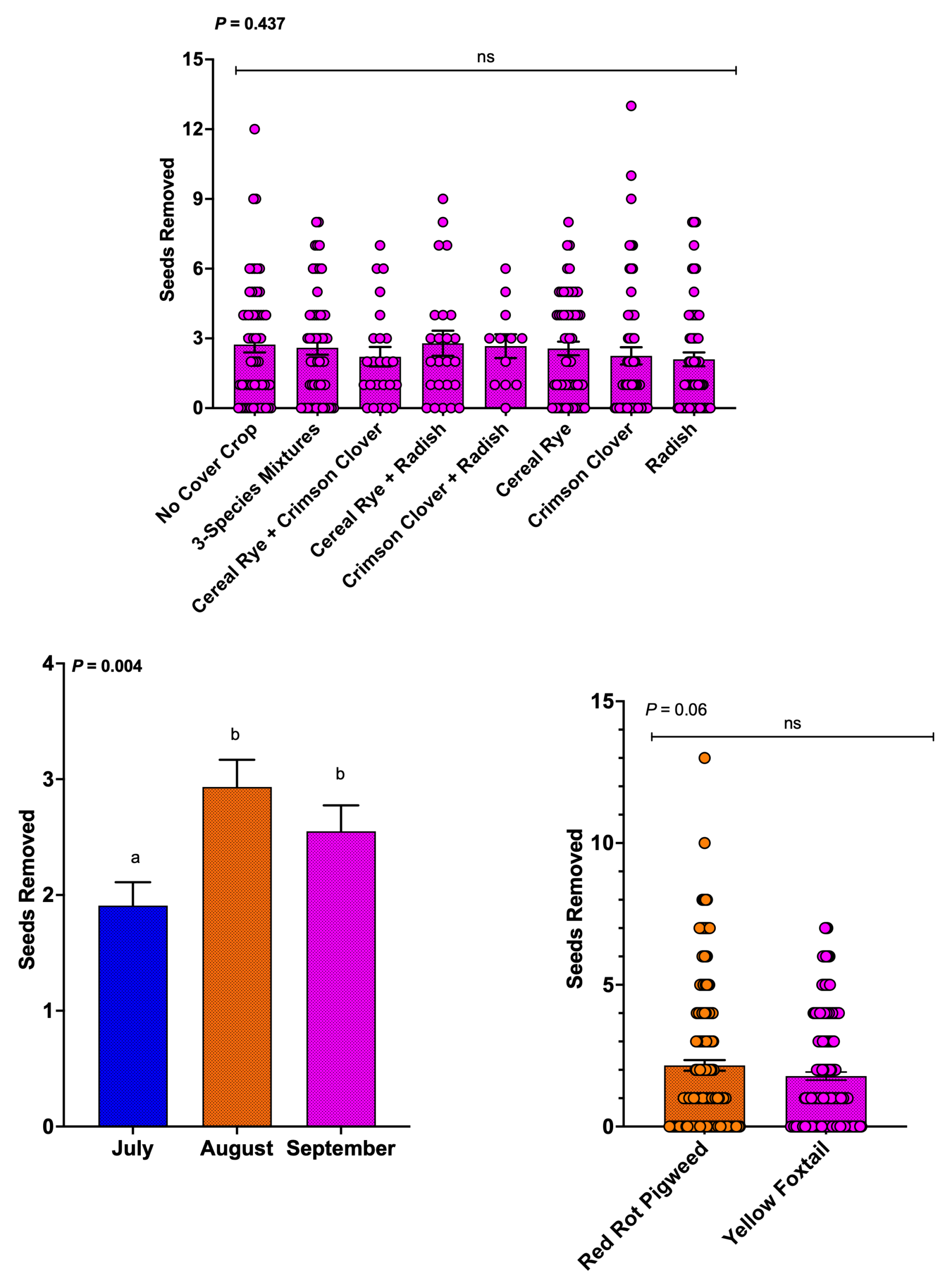
Predation of weed seeds
Cover crop treatment did not affect weed seed removal (P = 0.437). All treatments showed similar levels of seed predation. This lack of a treatment effect was unexpected, as we hypothesized that cover crops supporting higher ant abundance would also support greater seed removal.
Month significantly affected seed predation (P = 0.004). Seed removal increased in August and September relative to July. This seasonal pattern mirrors increases in ant foraging activity and predation across growing season, suggesting that greater predator activity in late summer contributes to higher weed seed removal.
Weed species did not affect seed removal rates (P = 0.06). Redroot pigweed and yellow foxtail seeds were removed at similar rates, although redroot pigweed showed a slight trend toward higher removal. These results suggest that predators consume weed seeds relatively indiscriminately, with potential implications for broad-spectrum weed management.
Late-season weed biomass
Cover crop treatment significantly affected late-season weed biomass (Gamma GLMM; P < 0.001). Crimson clover alone supported the highest weed biomass and significantly differed from cereal rye alone, which supported the lowest weed biomass. The three-species mixture, cereal rye + crimson clover, cereal rye + radish, and crimson clover + radish all supported low weed biomass comparable to cereal rye alone. Radish alone and the no-cover control supported intermediate weed biomass.
Contrary to expectations, crimson clover alone provided poor weed suppression. In contrast, cereal rye and treatments containing cereal rye consistently suppressed weeds. These results indicate that cereal rye provides strong weed suppression, likely through physical residue that persists into the cash-crop phase.
Treatments with lower weed biomass, including cereal rye and the three-species mixture, also supported higher ant abundance and waxworm predation. However, cover crop treatment did not affect weed seed predation, indicating that weed suppression in this system likely arises from direct cover-crop effects such as residue and competition rather than predator-mediated seed predation.
Species Richness
Cover crop treatment did not affect ant species richness (Negative Binomial GLMM; P = 0.403). All treatments supported similar numbers of ant genera, typically ranging from two to three genera per sample. This absence of a treatment effect was unexpected, as we hypothesized that cover crops providing greater habitat complexity or resource availability would support higher ant diversity.
Month significantly affected ant species richness (P < 0.001). Richness was higher in September than in July. This seasonal increase suggests that ant communities become more diverse as the growing season progresses, potentially due to colony establishment, increased foraging activity, or changes in habitat structure.
The contrast between significant treatment effects on ant abundance and the lack of treatment effects on species richness indicates that cover crops likely increase population sizes of species already present rather than increasing the number of ant species present in the system.
Shannon-Wiener diversity
Cover crop treatment significantly affected Shannon–Wiener diversity of ant communities (Linear Mixed Model; P = 0.006). Cereal rye + crimson clover and three-species mixture supported the highest diversity and differed from the no-cover control, which supported the lowest diversity. Most other cover crop treatments supported intermediate diversity. These patterns indicate that cover crop mixtures, particularly those that include cereal rye and legumes, promote more ant communities than the no-cover control.
Month also affected Shannon–Wiener diversity (P < 0.001). Diversity was higher in September than in July, mirroring patterns observed for species richness. This seasonal increase indicates that ant communities become both more species-rich and more evenly distributed as the growing season progresses.
The presence of treatment effects on Shannon–Wiener diversity but not on species richness suggests that cover crops influence the relative abundances of ant species rather than the number of species present. Treatments with higher diversity indices therefore supported more balanced ant communities rather than dominance by a single abundant species.
Simpson diversity
Cover crop treatment did not affect Simpson diversity of ant communities (Linear Mixed Model; P = 0.236). All treatments supported similar diversity values, with substantial variation within treatments. This result contrasts with the Shannon–Wiener diversity analysis, which showed significant treatment effects.
Month significantly affected Simpson diversity (P < 0.001). Diversity was higher in September than in July, consistent with patterns observed for species richness and Shannon–Wiener diversity. This consistent seasonal response across multiple diversity metrics indicates that ant communities become more diverse and more evenly structured as the growing season progresses.
Education & outreach activities and participation summary
Participation summary:
I developed a comprehensive course module on Ants in Integrated Pest Management for high school agricultural science teachers. This module is now available on the Global Teach Ag Network (GTAN), providing educators across multiple countries with resources to teach students about the role of ants in sustainable agriculture. Through a partnership with a teacher in Millerstown, PA, the module is already being implemented in the classroom, reaching high school students with hands-on lessons about ant ecology and IPM strategies.
To support broader dissemination of this research, I recently submitted a review paper on the functional roles of ants in no-till cropping systems for publication in Pest Management Science journal. This manuscript synthesizes current knowledge on ant ecosystem services and identifies opportunities for integrating ant conservation into farming practices.
To maximize the impact of my research results, I will engage in various educational and outreach activities. My advisor, John Tooker, has a 50% extension appointment and has established connections with field crop growers, agricultural professionals, industry representatives, and extension personnel throughout Pennsylvania, the Mid-Atlantic region, and the northeastern U.S. This network provides me with valuable opportunities to share my findings with the farming community during extension events and summer field days. The role of cover crops for promoting ant communities and their potential for sustainable pest control is a topic well-suited for Penn State events such as IPM Field Day, Farming for Success Field Day, Diagnostic Clinic, and many county-based events such as Crop Walks and Twilight Tours
I will also collaborate with organizations focused on soil health and conservation practices, such as the NRCS and the Pennsylvania No-Till Alliance, to explore opportunities for contributing to their events. These platforms will allow me to discuss benefits of no-till farming, cover cropping, and IPM for fostering beneficial insect communities, including ants, and their role in sustainable pest management. To ensure that the findings reach a wider audience, particularly field crop farmers and extension educators, I will collaborate with my advisor to create extension fact sheets and newsletters that summarize the key takeaways from my research.
In addition to agricultural audiences, I will also present my research results at the annual meetings of the Entomological Society of America, and regional Cover Crop Councils. I will also strive to publish my results in peer-reviewed journals.
Engaging younger audiences is equally important, and I plan to develop an interactive activity for Penn State’s Great Insect Fair. This activity will target children aged 5-12 and will focus on the fascinating world of ants in agroecosystems, emphasizing their role in pest control. I will also explore opportunities to educate high school students through programs organized by Penn State, such as Pennsylvania Governor’s School for the Agricultural Sciences, or for high school agricultural teachers, who can then share the information with their students. By sharing my passion for entomology and my research experiences, I hope to encourage these young individuals to consider careers in science and agriculture.
By implementing this comprehensive outreach plan, I will disseminate the findings of my research to a diverse range of audiences, including farmers, extension personnel, agricultural professionals, students, and the general public. This multi-faceted approach will contribute to a better understanding of the importance of ants in agroecosystems and the potential benefits of cover crop mixtures and no-till farming for sustainable pest management.