Final Report for GS05-049
Project Information
A comparison of various organic mulches in conjunction with winter and summer high biomass cover crops during herbicide-free no-till collard production found that mulches applied in the fall were effective in altering weed species composition away from broadleaves and sedges, although forage soybean as a summer cover crop was not, due to rapid decomposition. Persistence and nutrient release rates from organic residues result in slowly released nitrogen that may be available to subsequent crops. This study shows that it is possible to increase soil organic matter and nitrogen content while simultaneously keeping land productive.
Introduction
Organic agriculture is one of the fastest growing sectors in the US agricultural industry. The sale of organic foods in the United States had an average annual growth rate of 19.5% from 1998-2003, representing $10.38 billion in consumer sales in 2003 (OTA, 2004). Certified organic cropland in the US increased 103% between 1997 and 2005, while total acreage increase 201% (USDA, 2009). Although organic producers make efforts to adopt a sustainable approach on their farms, they often use conventional tillage because they cannot use herbicides to kill weeds and cover crops. Conventional tillage destroys organic matter, increases erosion risk, damages soil structure, reduces aggregate stability, promotes crusting and decreases soil moisture compared to no-till (Raczkowski et al., 2002; Bessam and Mrabet, 2003). Research conducted by Dr. Ron Morse has demonstrated the feasibility of organic no-till in Virginia (Rodale, 2005).
The key to organic no-till is the production of high biomass cover crops. High biomass cover crops are desirable because they contribute substantial amounts soil organic matter (SOM) and cover a large percentage of the soil surface. However, high biomass cover crops with low C/N ratios such as sunn hemp (Crotalaria juncea L.) require additional weed control as residues decompose (Ron Morse, personal commmunication, May 17, 2005), necessitating some form of weed control later in the season. On the organic farm, further weed control is often accomplished by hoeing, cultivating or purchasing off-farm inputs, which can be costly in terms of labor, transportation and input costs. Alternatively, mulching materials can be provided in situ by the production of high biomass perennial legumes grown as hedgerows in the field border or within the field itself, such as in alley cropping systems (Jordan, 2004). High biomass perennial legumes are desirable because they can provide a high quality mulch, rich in nitrogen (N) as well as other nutrients that become available to the crop as it decomposes, reducing the need to supply nutrients from other sources. Nutrients not used by the concurrent crop become potentially available to subsequent crops upon mineralization. Perennial legumes do not require re-seeding and often are more productive than annual species. By growing the legume mulch in the field, the costs of purchasing and transporting mulch are eliminated and the cost of providing nutrients to the crop is also reduced.
The rate at which plant residues and mulches decompose is dependent upon several factors. Mulches with a lower C/N ratio tend to decompose faster than those with a low N content, while presence of lignin and polyphenols slows decomposition rates (Fox et al., 1990). A balance must therefore be achieved between nutrient release and mulch persistence. Mimosa (Albizia julibrissin), a leguminous tree, and lespedeza (Lespedeza sericea) may be appropriate perennial leguminous species for mulch production because they can tolerate heavy pruning and have been shown to produce up to 4.0 U.S. tons of dry matter per acre in Alabama (Mosjidis, 1996; Kang et al., 2008). Although both species have a low C/N ratio, lespedeza is likely to decompose more slowly due to higher tannin concentrations, and thus persist longer as mulch (Kalburtji et al., 1999). Straw, a common organic mulch having a high C/N ratio, is likely to have slower nutrient release rates than mimosa prunings or lespedeza cuttings, and therefore may be more effective for weed suppression later in the growing season.
Summer cover crops have the capacity to improve soil quality, recycle or contribute nutrients, reduce weed growth, minimize soil erosion and produce large amounts of biomass in a short period of time (Creamer and Baldwin, 2000). After termination, nutrients released from residues are available for subsequent crops upon mineralization. Forage soybean (Glycine max) is an annual legume that may be a useful in the Southeast for its utility as a high biomass, low C/N summer cover crop.
Timing of cover crop termination is important in organic no-till systems. Crimping and rolling cereal cover crops may not be sufficient to produce an adequate kill if the crop is not mature. At the soft-dough stage of cereal growth, roller-crimping was as effective as herbicide in achieving an adequate kill (Ashford and Reeves, 2003). This period is identified as the 11.1 to 11.2 stage according to the Feekes Growth Stages (Large, 1954). This stage of growth is achieved around April 15-20 for rye (Donald Ball, personal communication May 23, 2005) in central Alabama. Alternatively, adequate termination of cover crops has been achieved by flail-mowing (Morse, 1999).
The objectives of our experiment were:
1. To compare mimosa prunings, lespedeza cuttings and straw as organic mulches in a high residue no-till system; and
2. To evaluate forage soybean as high-residue summer cover crop for no-till production of collards and the interaction of cover crop and mulch type on assessment parameters.
Mulches and cover crops were evaluated in terms of weed control over three years, decomposition and nutrient release rates, and crop yields.
Cooperators
Research
Weed suppression studies were conducted at the E.V. Smith Research Center in near Tallassee, AL (N 32º29.29’ W85º53.26, 66 m elevation) between 2005 and 2008 on a Wickham fine sandy loam soil, 0-2% slopes (Wickham fine-loamy, mixed, semiactive, thermic Typic Hapludults). The experiment was a 2 by 4 factorial randomized complete block design replicated four times. Each block was 24.4 m long and 9.1 m wide, with experimental units measuring 9.1 m long and 3.0 m wide. Two main treatments consisted of a Derry forage soybean (Glycine max (L.) Merr. cv. Derry, group VI) summer cover crop and a no summer cover crop control. Four sub-treatments consisted of in situ organic mulches: mimosa prunings ≤1 cm in diameter, lespedeza (Lespedeza sericea cv. AU Grazer) cuttings, wheat straw (Triticum aestivum L.), and a no-mulch control.
The plots were disk harrowed at the initiation of the experiment in October 2005, then limed and fertilized according to soil test recommendations. Each year, a winter cover of rye (Secale cereale L. cv. Elbon) was mechanically terminated using a roller-crimper (Ashford and Reeves, 2003) or chemically terminated if an adequate kill was not obtained in late April. Two weeks after termination, summer cover crop treatments were planted using Derry forage soybean at 112 kg/ha on 19 cm rows using a Marliss no-till drill. In mid to late August, summer cover crops were mechanically terminated using a roller-crimper or chemically terminated if an adequate kill was not obtained. Two weeks after summer cover crop termination, rows were cleared using row cleaners on a Kenzie no-till planter and collards cv. Champion seedlings were transplanted 43 cm apart using a single row RJV 600 no-till transplanter (R J Equipment, Ontario Canada) on 76 cm rows. No subsoiling shank was used at any point during the experiment. Mulches were applied at a rate of 6.7 Mg/ha (oven-dry basis) 21 days after transplanting. Collards were fertilized at a rate of 202 kg N/ha in three split applications and irrigated using a traveling gun as needed. Hand harvest operations were conducted 65-69 d after transplanting, followed by planting a winter cover crop of rye at a rate of 101 kg seed/ha on 19 cm rows. Weed coverage was determined using line-transect methodology by counting 50 points along a marked line twice per plot per sampling period. Weeds were classified as broadleaves, grasses or sedges. Twice during 2008, weeds were identified to the species level. Yield was determined by hand-harvesting two 2-m rows within the sampling area. The harvested heads were counted for marketable heads and weighed.
Significant effects were identified by analyses of variance as implemented in SAS 9.1.3 using PROC GLIMMIX procedures and maintaining blocks as a random effect (SAS, 2003). Reduced models were obtained via backward elimination for variable selection using P > 0.15 as the criteria for elimination from the model. Inflated Type I error rates associated with the covariance structure in the model was limited by adjusting the denominator degrees of freedom using Kenward-Roger correction in the MODEL statement (Littell et al., 2002). Means and standard errors of significant effects of the reduced models were obtained using PROC MEANS.
A field decomposition study was conducted at the same location using four organic residues. Lespedeza, mimosa, oat (Avena sativa) straw, and soybean (Glycine max var. Stonewall, group VII) were obtained locally to supply residue. Air-dried residues were packed into nylon mesh bags measuring 20 cm by 10 cm with 50 to 60 micron openings at a rate equivalent to 6.7 Mg/ha (3.0 tons/ac) on an air-dry basis.
Sealed litterbags were placed on the soil surface or buried at 10 cm depth on Oct. 9, 2007. The site was maintained under no-till for three years prior to placement. Conventional till plots were disked immediately before placement. The treatments were arranged in a randomized split-plot design with four replicates. Bags were retrieved from the field periodically at 0, 3.5, 7, 14, 28, 56, 112, and 224 days after application. The contents of each bag were oven-dried and weighed for dry matter determination. They were then ground to pass a 16 mesh sieve and analyzed for total C and N by LECO TruSpec CN (Leco Corp, St. Joseph, MI). Sample contamination by soil was accounted for by converting all data to an ash-free dry weight basis by ashing approximately 1 g of the samples in muffle furnace at 400oC for 12 h and determining the ash free dry weight (Cochran, 1991).
Means, standard errors, and statistical significance of treatments were determined using Proc Mixed (SAS, 2003) at the 95% confidence level. Least squares estimates for nonlinear models were determined using four parameter double exponential decay models (Systat Software Inc., 2006).
Weed suppression.
Mulching provided weed suppression of broadleaves, grasses and sedges, but forage soybean as a summer cover crop did not (Table 1), likely due to the fact that soybean residue decomposes too quickly to have a lasting mulching effect. In all cases, days after mulching (DAM) were highly significant within each year of the study (P < 0.0001). A time by mulch interaction within each year (DAM*Mulch(Year)) was due to both the effect of mulch application and the growth of weeds as the season progressed. Evidence of a mulch by year interaction suggested that weed populations were affected by mulching for three consecutive years. This effect was most apparent on broadleaf control (Figure 1). Mulching the first year was effective for suppression of broadleaf weeds. Suppression of broadleaf weeds during the first year appears to lessen broadleaf infestation during subsequent years, although mulching during 2007-8 did not have the same level of suppression compared to the non-mulched control. Since conservation tillage tends to shift weed populations away from broadleaves (Teasdale et al., 1991; El Titi, 2003), it is not surprising that broadleaf control can be enhanced with mulch application.
| Effect | P > F | ||
| Broadleaf | Grass | Sedge | |
| Mulch | 0.0913 |
0.0315 |
0.0043 |
| Mulch*Year |
0.0054 |
0.1077 | 0.1046 |
| DAM(Year) | <.0001 | <.0001 | <.0001 |
| DAM*Multch (Year) | 0.1128 | 0.0014 | 0.0008 |
| Year | n/s | n/s | 0.0154 |
| CC*Mulch | n/s | n/s | 0.0924 |
| CC*Mulch*Year | n/s | n/s | 0.1074 |
Table 1. Probability of greater F values for the effect of mulch, cover crop (CC), days after mulching (DAM) and year on weed coverage. Treatments not shown or not significant (n/s) were excluded after backward elimination variable selection for the reduced model if P > 0.15.
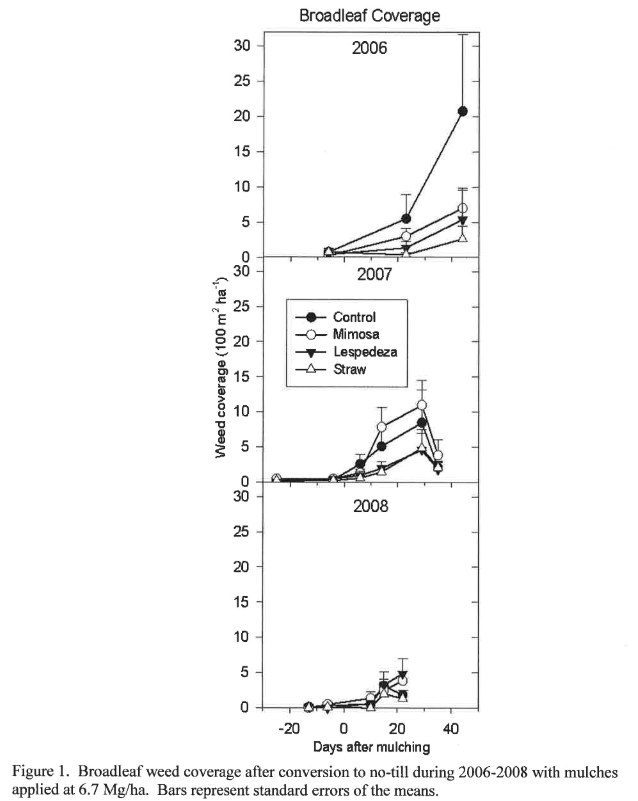
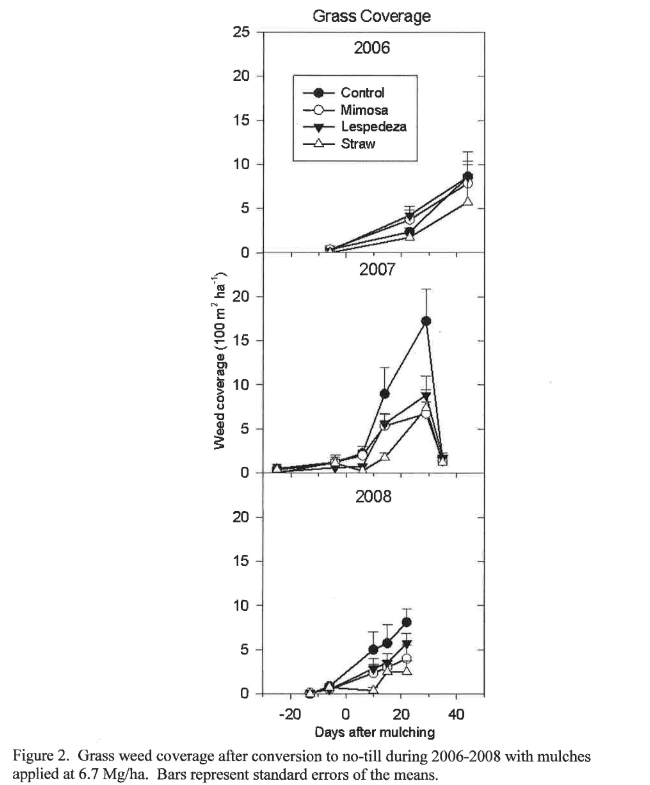
The population shift toward grasses under conservation tillage makes grass control more difficult (El Titi, 2003). During the first year of no-till, grass control was not effective using any of the mulches (Figure 2), but subsequent years showed much better grass suppression using mulches compared to the non-mulched control. Grass infestation was maintained below 10% by the application of any mulching material in 2007 (compared to 17% for the non-mulched control), and below 6% in 2008. The data show that mulching suppresses monocot weed populations in no-till systems if used for more than one year compared to the control, and may suggest that >2 years of no-till with high-biomass producing cover crops may be effective at reducing grassy weeds, although more data are needed to support this claim. In any case, the data show that grass populations under no-till are highly variable, with populations increasing dramatically during the second year of conversion from conventional tillage, but decreasing in the third year. Mowing grasses before seed heads become viable may reduce the grass populations to manageable levels during transition to conservation tillage.
During the first year of transition from conventional to conservation tillage, yellow nutsedge (Cyperus esculentus L.) control was highly problematic, with total coverage ranging from 7-21% (Figure 3). However, subsequent years of high residue no-till improved sedge suppression, generally below 5% coverage, although there was not much difference between any of the mulching treatments and the control. Bangarwa, et al. (2008) showed that straw mulch applied at 7300 kg/ha (7 cm thick) was effective at reducing medium (0.26-0.50 g) purple nutsedge (Cyperus rotundus L.) tuber density, but did not reduce large (>0.50 g) or small (0.10-0.25 g) tuber density for bell pepper production in Clemson, SC. They also found generally comparable tuber density when tilled plots were either straw-mulched at transplanting or hand-weeded every 1-2 weeks. It is interesting to note that sedge coverage was subject to a significant cover crop by mulch interaction (Table 1), resulting from increased sedge suppression by mimosa prunings after a forage soybean summer cover crop in 2006 and increased sedge suppression achieved in control plots when combined with forage soybean in 2007 and 2008 (Figure 3). Although it is unclear why this should be the case, it is apparent that sedge suppression is improved during subsequent years of high-biomass producing cover crops in combination with no-till, with or without the application of mulches. Yellow nutsedge was the only perennial weed species present after three years (Fig. 4).
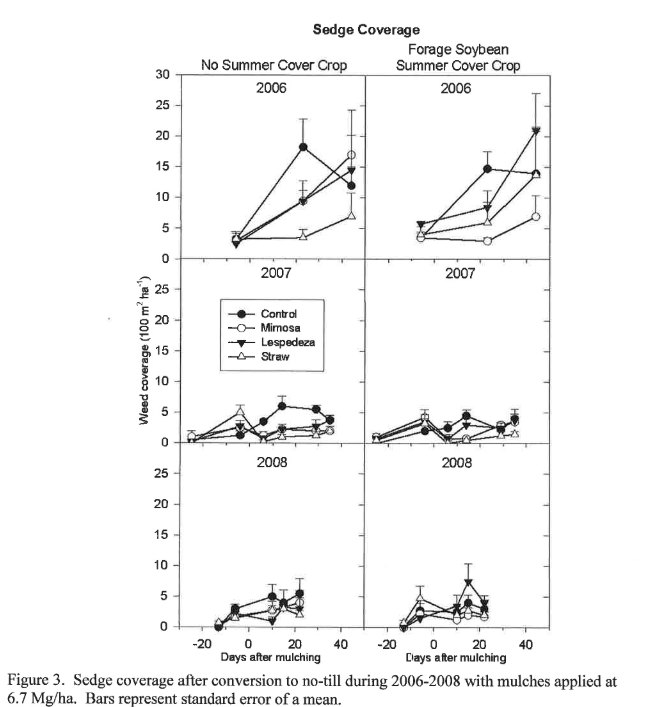
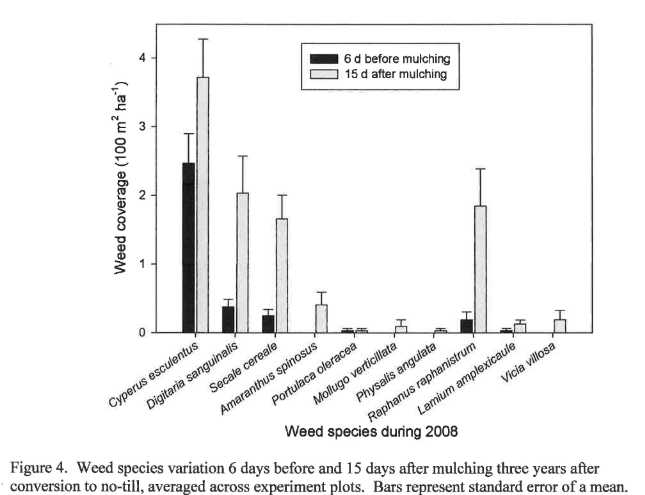
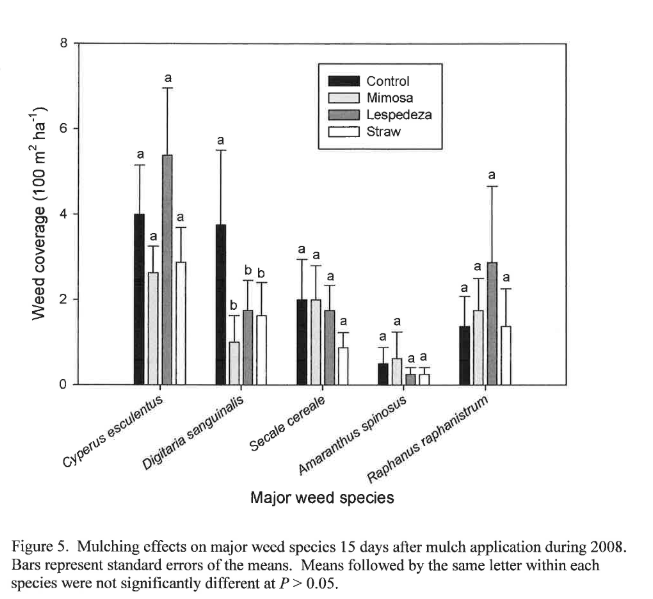
Figure 4 shows the weed infestation 6 d before and 15 d after mulching in 2008 by species averaged over all plots, giving an overall picture of the weeds present three years after initiation of no-till. No individual weed species covers more than 4% of the surface 15 days after mulching, or five weeks after transplanting. This suppression should give the main crop an adequate start to compete successfully with weeds later in the season. Yellow nutsedge was the single major species present at that time, followed by large crabgrass. After three years of high-biomass no-till, the only grasses present were large crabgrass and winter rye, the latter due to viable seed germination from the previous winter cover crop. This underscores the importance of ensuring termination of cover crops and mulches before seeds become viable. Summer annual broadleaves consisted of spiny pigweed (Amaranthus spinosus L.), common purslane (Portulaca oleracea L.), carpetweed (Mollugo verticillata L.) and cutleaf groundcherry (Physalis angulata L.), though all of these weeds were under very good control three years after initiation of no-till. It may be that these were not a major problem because experiment occurred in the fall or because of residual allelopathy from the winter rye. Burgos and Talbert (1996) found that rye, wheat, and rye with hairy vetch suppressed 70-85% of redroot pigweed (Amaranthus retroflexus L.) and yellow nutsedge eight weeks after cover crop termination without herbicides, and that rye alone and rye with vetch suppressed 65-70% of large crabgrass (Digitaria sanguinalis (L.) Scop.).
Among the winter annual broadleaf weeds, wild radish (Raphanus raphanistrum L.) exhibited much greater average groundcover than henbit (Lamium amplexicaule L.), though the average coverage was still less than 2%. Even so, fall mulching was not effective for wild radish control 15 d after application (P=0.6738) in 2008 (Figure 5). The same can be said for all the major weed species present 15 days after mulching during 2008 with the exception of large crabgrass. This may be due to the fact that weed coverage was already under considerably good control after three consecutive years of no-till with high-biomass cover crops, given that even non-mulched control plots exhibited less than 4% coverage of any particular species. The mat of residue on the soil surface after three years of no-till appears to be effective at weed suppression. Figure 5 also shows that while not statistically significant, straw mulch tends to be the best suppressor of the major weed species in 2008, likely due to the thickness of the straw residue.
Collard yield was not significantly different for any variable, including year (data not shown). This is not unexpected since crop yields are generally more responsive to tillage systems and management than to weed density alone (El Titi, 2003).
Persistence and decomposition rates.
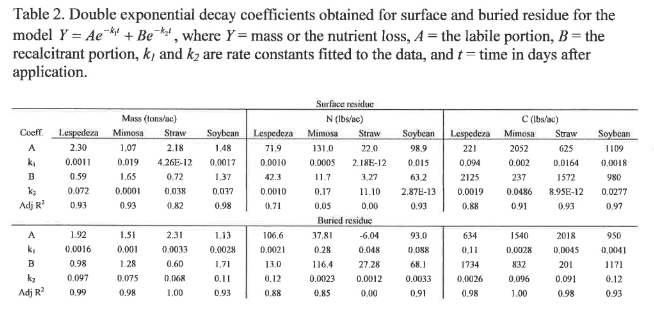
Decomposition of organic residue occurs in two phases. Initially, a labile portion of the residue, such as sugars, starches and proteins, is readily consumed by soil microbes, leaving behind a recalcitrant portion of the residue, such as cellulose, fats, waxes, lignin and tannins (Wieder and Lang, 1982). This recalcitrant portion is slowly decomposed and contributes to the development of organic matter in soil. Such a system is best described by double exponential decay models, with one exponential segment describing the labile portion and the other exponential segment describing the recalcitrant portion of the residue (Wieder and Lang, 1982). The double exponential decay model is represented by the equation Y = Ae^(-k1*t) + Be^(-k2*t), where Y = the nutrient or mass remaining, A = the labile portion, B = the recalcitrant portion, k1 and k2 are rate constants fitted to the data, and t = time in days after application. This model serves as the basis for comparison of N, C, and mass loss between conservation and conventional tillage in this study.
Mass loss from organic residues under conservation and conventional tillage is shown in Figure 6. Buried residue generally exhibits faster mass loss in both the labile and recalcitrant portions of all residues, as shown by the greater rate constants k1 and k2 for buried material compared to surface residue (Table 2).
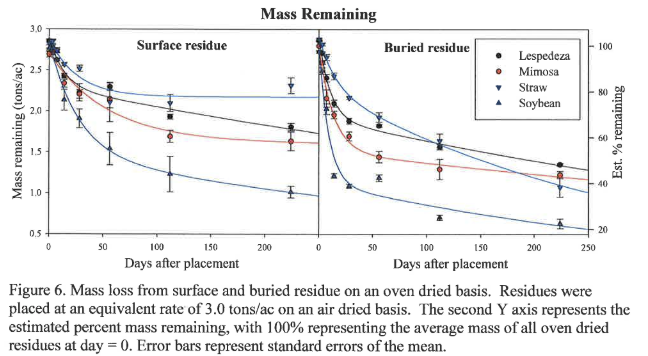
Nitrogen loss from organic residues under conservation and conventional tillage is shown in Figure 7. Buried residue generally exhibits faster N loss in both the labile and recalcitrant portions of all residues. This is evidenced by the greater rate constants k1 and k2 for buried material compared to surface residue (Table 2), though notable rate constant exceptions exist in cases where the curve fit (Adj. R^2) is exceptionally low, such as in the case of straw, which has a very low original N content and negligible labile N pool. For residues with a high N content, there is considerably more N potentially available to a spring crop from surface residue than buried residue. For example, at planting on May 1 (day 204), there is approximately 33 lbs/ac more N potentially available from surface soybean residue than incorporated soybean residue. Upon mineralization, N is subject to the competing processes of nitrification, immobilization, plant uptake, ammonium fixation, and volatilization. This study does not determine the fate of the lost N (i.e., the proportion mineralized, immobilized, etc.).
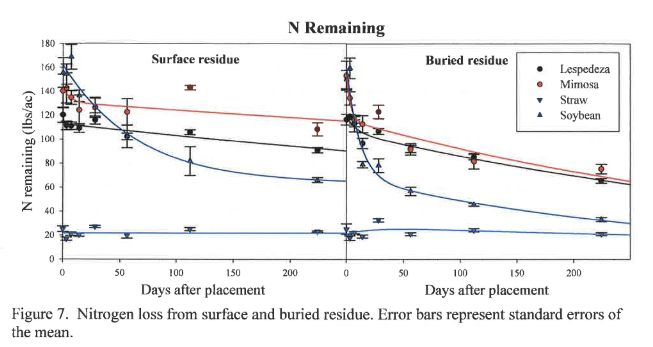
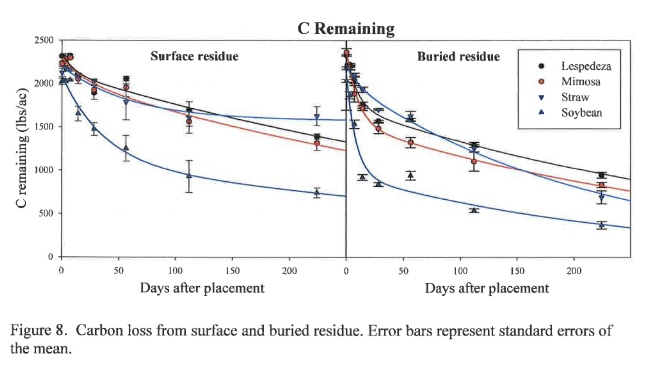
Carbon loss from organic residues under conservation and conventional tillage is shown in Figure 8. Buried C loss models appear similar to buried mass loss models (Figure 6) because most mass loss is due to the respiration of C, which is then lost to the environment as CO2. Buried residue exhibits faster C loss in both the labile and recalcitrant portions of all residues, as shown by the greater rate constants k1 and k2 for buried material compared to surface residue (Table 2). Carbon is therefore sequestered longer when residue is left on the surface compared to residue incorporation. This should result in greater SOM accumulation from surface residue over time. On a more speculative note, in an age when producers may be compelled to participate in a C market, conservation tillage practices may provide producers with a C offset or credit, while also enhancing SOM.
Educational & Outreach Activities
Participation summary:
Mulvaney, M.J., C.W. Wood, K.S. Balkcom, J. Kemble, and D.A. Shannon. 2017. No-till with high biomass cover crops and invasive legume mulches increased total soil carbon after three years of collard production. Agroecology and Sustainable Food Systems 41: 30-45. doi:10.1080/21683565.2016.1236766.
Mulvaney, M.J., A.J. Price, and C.W. Wood. 2011. Cover crop residue and organic mulches provide weed control during limited-input no-till collard production. J. Sustainable Agric. 35: 312-328. doi:10.1080/10440046.2011.554315.
Mulvaney, M.J., A.J. Price, C.W. Wood, and J.M. Kemble. 2009. Cover crop residue and organic mulches provide herbicide-free weed control during no-till collard production. Currently under review by the Journal of Sustainable Agriculture.
Mulvaney, M.J., C.W. Wood, and B.H. Wood. 2009. Persistence and nutrient release from organic residue under conservation tillage. 19th GSC Research Forum, Auburn, AL. Mar. 11-12, 2009.
Mulvaney, M.J., C.W. Wood, and B.H. Wood. 2009. Persistence and nutrient release from organic mulches and cover crops. 106th Southern Assoc. of Agricultural Scientists, Atlanta, GA. Feb. 1-3, 2009.
Mulvaney, M.J., C.W. Wood, B.H. Wood, K.S. Balkcom, J. Kemble, D. Shannon. 2008. High biomass cover crops and organic mulch effects on soil moisture and weed distribution during collard production. ASA-CSSA-SSSA Joint National Conference, Houston, TX. Oct. 5-9, 2008.
Mulvaney, M.J., C.W. Wood, and B. Wood. 2008. Nutrient release rates from organic mulches and cover crops. Proc. 8th Annual Conservation Production Systems and Training Conference, Tifton, GA. July 29-31, 2008.
Project Outcomes
Although mulching and summer cover crops did not have a direct impact on collard yield, mulching in conjunction with high biomass cover crops did provide substantial suppression of weeds, particularly broadleaves and sedges. Mulching over a number of years tends to shift weed populations rather than eliminate them. Weed populations were highly variable, with broadleaf and sedge populations decreasing over three years under the conditions of this study. Furthermore, mulching at a rate of 3.0 tons/ac provides a reasonable level of grass control under continuous no-till. Although main crop yields were not affected by application of various organic residues within the first three years of no-till, it seems likely that application of these residues will improve soil quality over time while simultaneously limiting external inputs.
In addition, mulches left on the surface increase soil organic carbon content, which will improve SOM status over time. However, producers should be aware that increased SOM content will not be evident after a single season or even a single year. Our research implies that tillage can destroy the accumulated SOM content simply by burying the residue, which aerates and warms the soil, and increases the contact of organic matter with the soil microbial community, with the result of soil microbes metabolizing the accumulated organic matter and respiring it as carbon dioxide. All producers, but especially organic producers, would benefit from retaining C as SOM instead of having it respired as CO2, thus the incentive for organic producers to adopt conservation tillage.
Buried residues decompose faster and release C and N quicker than surface residues. A winter cover crop may be able to recapture some of the N lost from buried residues in order to make it potentially available to spring crops. However, surface residues with a high N content retain N longer, and may provide more potentially available N to spring crops than buried residues. As such, surface residues may act as a slow release N fertilizer and contribute to organic matter accumulation on the soil surface. This study demonstrates that in situ cover crops and mulches may be utilized for the enhancement of SOM and soil N status.
Farmer Adoption
Producers considering the adoption of conservation tillage may wish to consider the use of high biomass cover crops in conjunction with organic mulches in order to suppress weeds and improve SOM content. Our research shows that it is possible to simultaneously obtain an annual harvest and improve SOM status in the Southeast. Some important factors that should be considered when applying this system include: 1. Utilize mulches that do not contain viable seeds. 2. Planting small-seeded cover crops into thick residue may be problematic, although the use of row-cleaners may circumvent this issue.
Areas needing additional study
Further studies need to be conducted to determine nutrient cycling efficiency, nutrient redistribution and release rates, organic matter accumulation, N mineralization rates from organic residue, and C sequestration during continuous high residue no-till with organic mulches. As agricultural sustainability becomes increasingly vital for political and food security around the globe, it is important that obstacles to sustainable food production systems be overcome.