Final Report for GS11-104
Project Information
This study uses native stinkbug egg parasites as a model to improve understanding of egg parasite overwintering and adult nutrition requirements, with a long-term goal of enhancing biocontrol of stinkbugs and their relatives (e.g. kudzu bug). Results show parasites benefit equally from honeydew (widely available on farms) as well as flower nectar. Field and lab studies indicate the importance of woodland field borders with hardwood leaf litter as winter refuges for egg parasites and kudzu bugs. This study should help to guide future conservation biocontrol efforts with both native and imported scelionid egg parasites of stinkbugs and their relatives.
Introduction
The purpose of this project is to improve management of stinkbug pests by focusing on conservation biological control of a parasitic insect, Telenomus podisi, that attacks stink bug eggs. The practice of conservation biological control seeks to modify aspects of agricultural environments to enhance beneficial organisms. Although insecticides are effective against stinkbug adults and nymphs, they have been found to be ineffective against eggs but lethal for egg parasitoids. Insecticides, whether they are synthetic or organic, can have significant impacts on beneficial insects and other ecological components in farm landscapes, as well as human applicators. As an external farm input, they are a recurring cost to growers, and reducing their need will improve agricultural sustainability. This study proposes to identify overwintering refuges that the egg parasitoid Telenomus podisi might utilize on farms as well as types of carbohydrate sources utilized for its daily functioning. This information will help us to provide recommendations to growers on what plants and trees to keep or maintain on farms to enhance on-farm populations of the parasitoid, and reduce potentially damaging populations of stink bugs. In addition, overwintering preference of the invasive pest kudzu bug, Megacopta cribraria (Hemiptera: Plataspidae), in relation to leaf litter and tree bark will be assessed.
1. Determine the presence of overwintering T. podisi in potential refuge sites like tree bark, dried fruiting and flowering bodies, insect cocoons, fallen pine cones etc. that might be utilized by T. podisi in the field.
2. Determine the lifespan and reproductive ability of T. podisi when offered carbohydrate sources in the form of various flowering plants, and homopteran honeydew.
Cooperators
Research
Objective One
In 2012 three new sites were targeted for winter sampling near Clayton (Johnston County, NC) that had a south-facing woodland border next to a soybean field with stink bug populations. A total of 58 fine mesh cages with a funnel-bottle collection system at their tops were wrapped around 3 feet of tree boles near the ground in January 2013 (Fig.1). Ten pairs of pine and sweet gum trees were caged at each of two locations, and 7 pines and 11 sweet gums were caged at the third location. Insects that emerged in the spring from tree bark were collected and identified. In addition to tree caging, leaf litter samples were collected from 1 sq. ft. areas adjacent to trees on the north and south sides of caged trees, and between tree pairs, at the first two of the three locations. These samples were taken to a greenhouse for extraction using mesh bags with funnel-bottle collectors at the top (Fig. 2). Kudzu bugs, Megacopta cribraria (Hemiptera: Plataspidae) and parasitoids were collected, preserved in 70% ethanol, and identified.
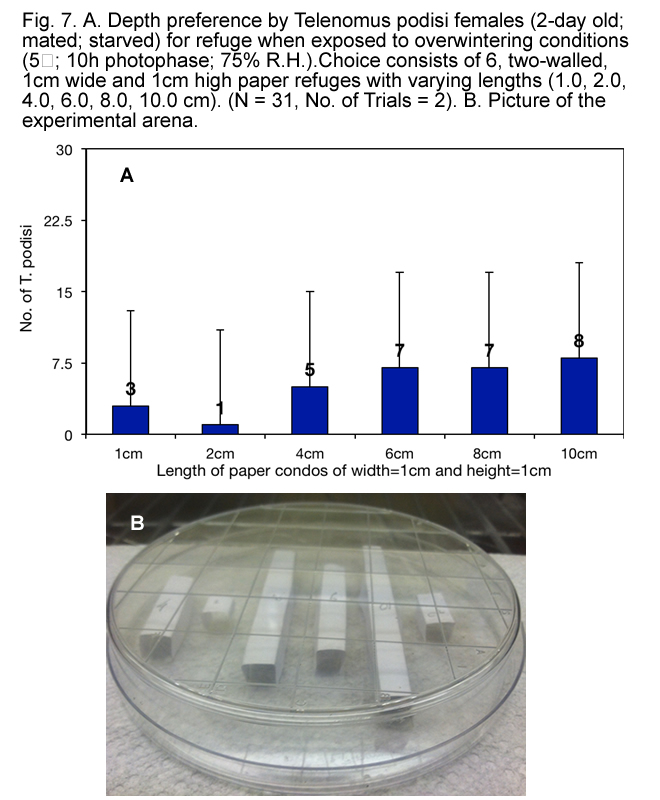
A series of laboratory studies were done to better understand the space and orientation requirements of T. podisi females under overwintering conditions. Refuges of varying dimensions were built out of paper or hatched stink bug eggs and presented to 2-day old mated T. podisi females in petri-dishes. These arenas were exposed to overwintering conditions over a 5 day period in a growth chamber starting at 25C and 14h photophase, then reducing the temperature 5C and photophase 2h each day to reach 5C and 10h photophase on day 5.
For each of the experiments, on day 5 the position of wasps was observed and reported. To ascertain that wasps were not being attracted to the glue used in making refuges, 2-day old mated females were presented with a 2 choice test wherein one refuge (dimensions of 1cm x 1cm x 1cm) was glued to parafilm, which in turn was glued to the base of the petri-dish, while the other simply rested on the base of the petri-dish, with no glue used on it. Results indicated that 29 of 50 wasps went inside either of the paper refuges. Of those 29, 15 went in the refuge with glue and 14 went in the refuge without glue, indicating no preference for refuges with glue. Additional experiments conducted on the wasps with different types of refugia were done with position of the refuges randomized within the petri-dish.
Data Analysis. Log of 1 + number of kudzu bugs overwintering in leaf litter transformation was done to achieve approximate normality and used for comparison of least square means. Analysis of Variance was followed by Fisher’s Protected LSD (P ? 0.05) (PROC GLM, SAS Institute, 2013) for separation of means testing.
Objective Two
Colonies of the following species of insects and plants were reared throughout 2012.
Insects: 1) Podisus maculiventris (Spined soldier bug); 2) Telenomus podisi (Egg parasitoid); 3) Tenebrio molitor (Mealworm/ Darkling Beettle) (Food source for the P. maculiventris colony); 4) Aphid species (Aphis craccivora). Plants: 1) Fagopyrum esculentum (Buckwheat) (for flowers to be used in Objective 2 studies); 2) Glycine max (Soybean) (for P. maculiventris colony); 3) Vicia fava (Fava Beans) (for aphids to be used in Objective 2 studies)
Longevity and fecundity of T. podisi was estimated in cages (Fig. 10) using a randomized complete block design, with position of cages on shelves in a rearing-room acting as blocks. The treatments were as follows: Treatment 1 - Exposure of a single 2-day old mated female T. podisi to flower of buckwheat still attached to the plant. Treatment 2 - Exposure of a single 2-day old mated female T. podisi to a single fava bean leaf infested with cowpea aphids. The leaf remained attached to the plant. Leaves selected were not too old (near the base of the plant) or too young (at the apex of the plant), but rather in the middle, so that they did not wilt easily and were large enough to support a respectable number of aphids. The leaf was allowed to have 5 aphids at the start of the experiment. Control 1 - Clover honey in single mated 2-days old female T. podisi trials was provided as a single streak of pure honey approx. 50 mm long applied with an insect pin to the internal wall of the cage on the first day of each replication. Control 2 - Organic clover honey in single mated 2-days old female T. podisi trials was provided as a single streak of pure honey approx. 50 mm long applied with an insect pin to the internal wall of the cage on the first day of each replication. All the abovementioned treatments and control cages were misted with tap water everyday. Control 3 - Only tap water was misted on the cages regularly. Experimental cages were fashioned according to Witting-Bissinger et al. (2008). All experiments using T. podisi were carried out in a rearing room at 25 ± 0.1 °C, 70 ± 1.3% RH, and a 14 h photophase. Potted plants were placed on metal shelves so that flowers can be inserted between slots in the shelf above. Flower heads were positioned so that the head touches the inside of the lower screen and side of cages. Plants and their flowers were replaced when approx. 50% of florets had senesced. A single leaf, infested with cowpea aphids were also positioned in the treatment cage similarly. The leaf was replaced when it started showing signs of wilting or drying. Longevity of the wasps was recorded.
Fecundity was assessed for T. podisi during the same experiment used to monitor longevity. Each female T. podisi was provided with one Podisus maculiventris egg mass per day. The number of eggs per mass was rationed with 20 eggs/mass on the first day of the experiment. In the next 3 days, 10 eggs/mass were provided daily. 5 eggs/mass were provided as long as the wasp survived from thereon. After every 24 hours, each egg mass was collected in vials and held for offspring emergence in the same rearing room. Only those females that produced both female and male progeny (i.e. mated) were included in data analysis.
Data Analysis. Calculation of means (±SD) was done using untransformed data for both longevity and fecundity experiments which were replicated 5 times. Data for organic honeydew fed T. podisi was obtained only during the last 2 replicates. All longevity data was log transformed to achieve approximate normality. Analysis of Variance was done on longevity and fecundity data to determine the difference in means. Fisher’s Protected LSD (P ? 0.05) (PROC GLM, SAS Institute, 2013) was used for separation of means.
{kind=link}
{kind=link}
Objective One
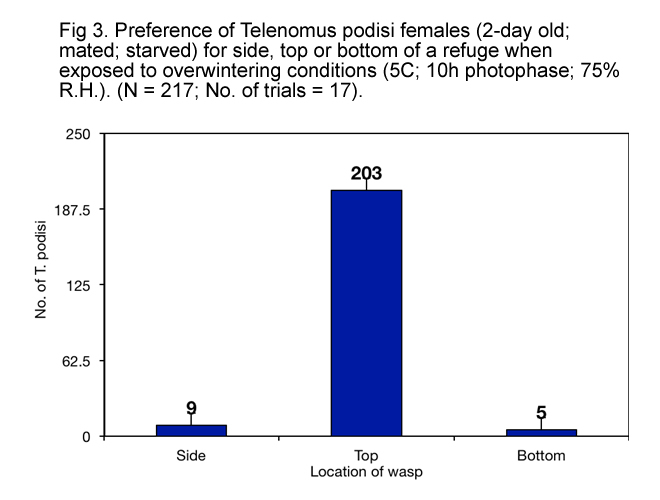
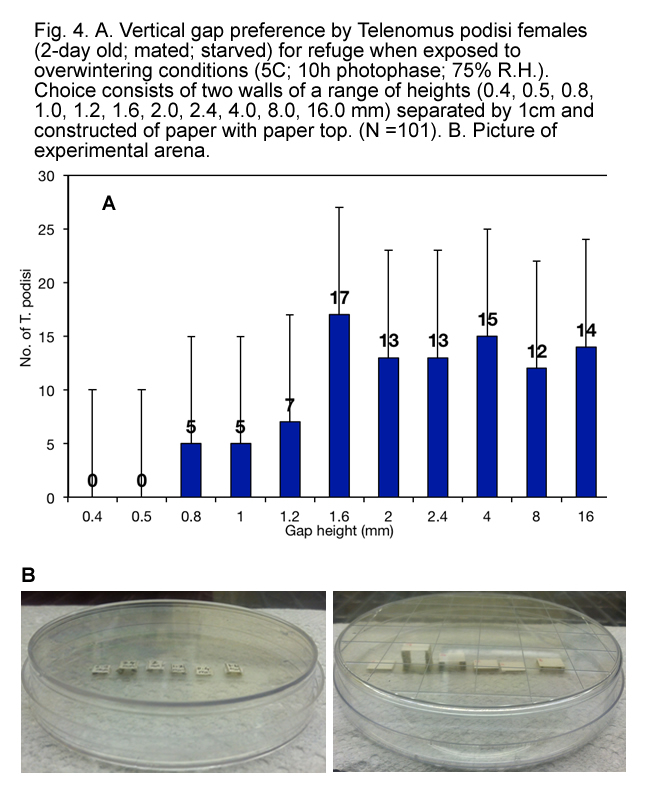
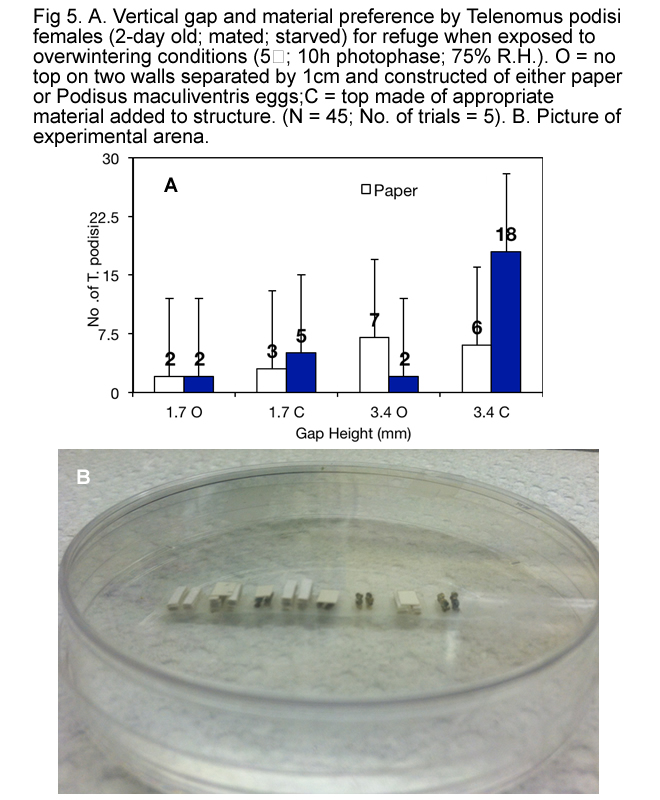
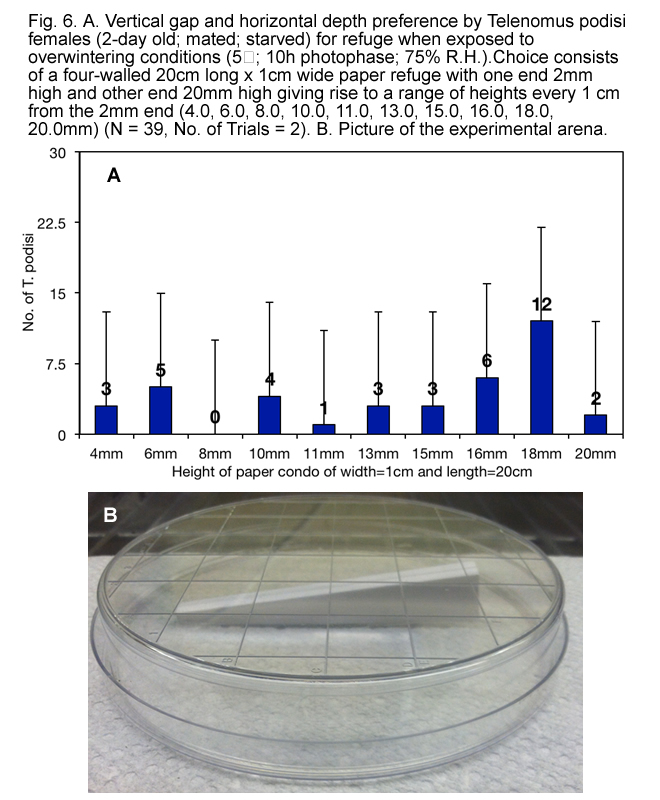
It appears that, when exposed to overwintering conditions, T. podisi preferred to hang upside down from tops of refuges made of natural materials (Fig. 3) that have at least 1.5 mm between surfaces (Fig.s 4, 5, 6 and 7), and preferred refuges that offered more cover. This suggested that T. podisi may prefer leaf litter as a winter refuge.
Kudzu bugs in leaf litter and tree bark
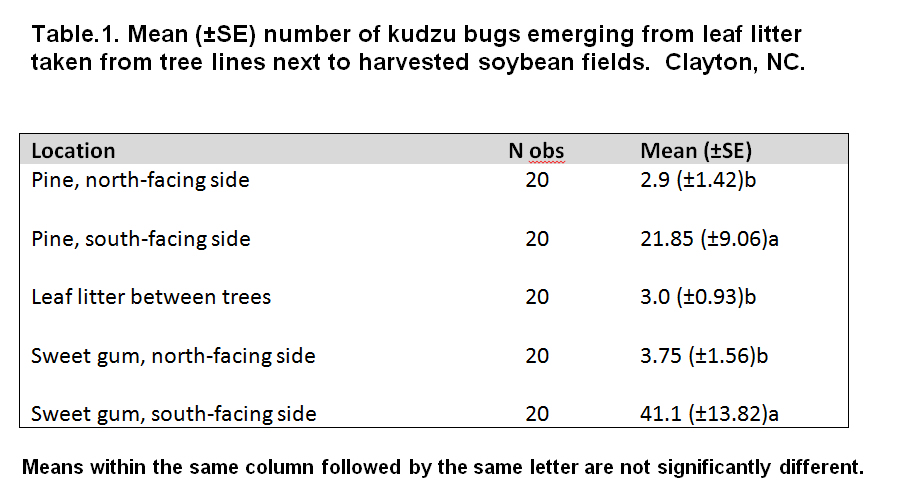
Location of the leaf litter was found to be a significant factor in determining the number of kudzu bugs overwintering in leaf litter (F = 6.56, df = 25, 74, P < 0.0001). Mean number of kudzu bugs found overwintering in south-face leaf litter of sweet gums was 1.88 times the same in south-face of pines (Table 1), 11.0 times the same found in north face of sweet gums, 14.2 times the same found in north face of pines and 13.7 times the same from between the tree pairs. Site and tree-pair effect was found to be strong (F = 2.85, df = 18, 74, P = 0.0008).
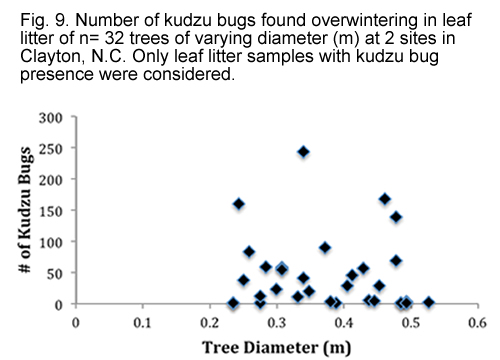
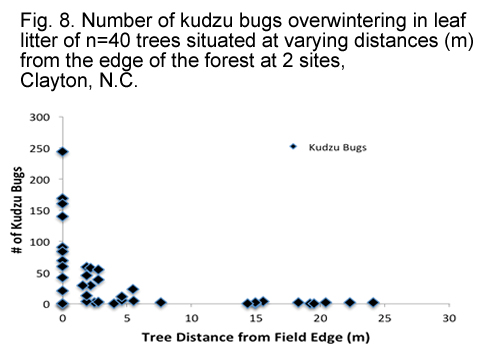
Distance of the leaf litter sample from the edge of the forest was found to be significant factor in influencing the number of overwintering kudzu bugs (F = 24.30, df = 1, 74, P < 0.0001). A strong quadratic distance effect was also seen (F = 10.87, df = 1, 74, P = 0.0015). Fig. 8. shows the distribution of overwintering kudzu bugs in leaf litter as the distance of the trees from the edge of the forest increases. However, diameter of the trees was found to have no effect (F = 0.23, df = 1, 55, P = 0.6327). Fig. 9. shows the distribution of overwintering kudzu bugs in leaf litter as the diameter of the tree increases. Mean tree diameter was 0.38 (± 0.09) meters.
No significant effect of location and site was seen on number of kudzu bugs overwintering in bark (P > 0.05). A small number of kudzu bugs were found overwintering in bark with a mean of 0.3 (± 0.13 S.E.) in pine trees and 0.1 (± 0.07 S.E.) in sweet gum.
Parasitic wasps in leaf litter and bark
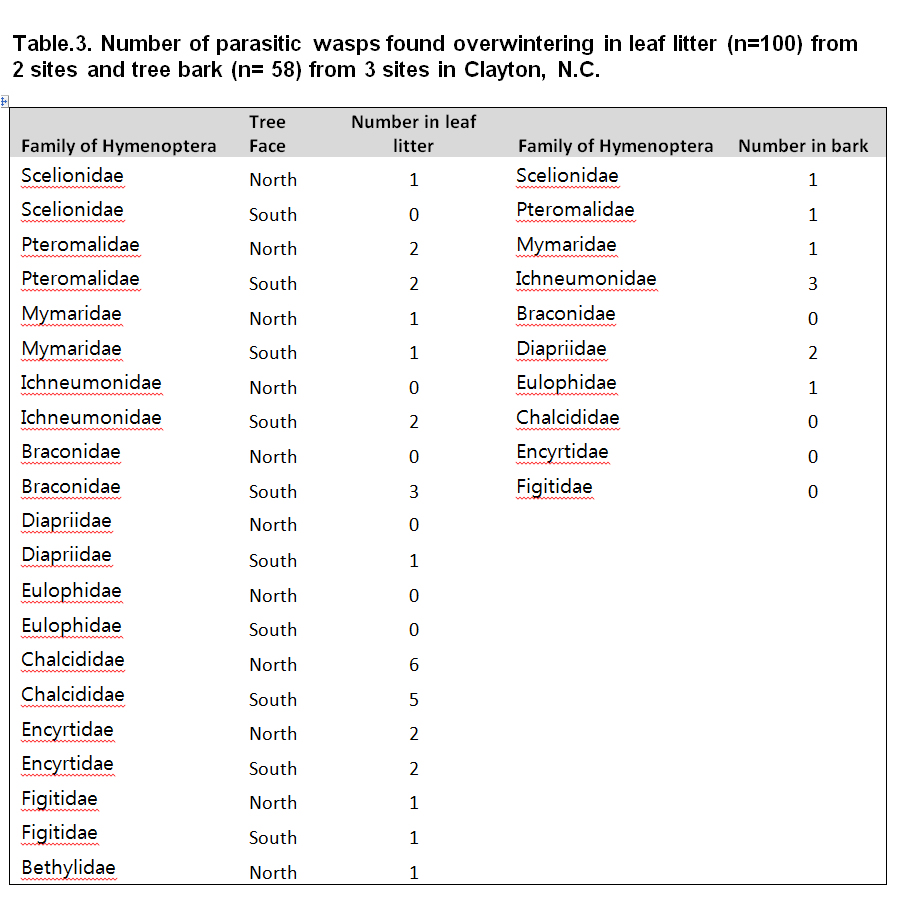
A total of 50 samples of leaf litter and 40 samples of tree bark from 2 sites yielded very low numbers of parasitic wasps compared to kudzu bugs. Therefore, no statistical analysis was possible (Table 2). However, it was found that the mean number of parasitic wasps overwintering in leaf litter was 4.3 times greater than those overwintering in tree bark. Table 3. shows the numbers of different families of wasps found overwintering in leaf litter and tree bark. Overwintering parasitoids were found in leaf litter at the rate of 0.31 /square meter.
From the results, it can be concluded that both parasitoids and kudzu bugs preferred to overwinter in leaf litter as opposed to tree bark. Also, it appears that kudzu bugs tend to remain closer to the edge of the forest rather than overwintering into deeper sections of the forest bordering the crop field. There was a clear preference of the south-facing leaf litter as overwintering site by kudzu bugs. Although, the derth of parasitoid data prevented the establishment of a similar trend, it has been seen that in parasitoids, the black body color helps the overwintering wasps to capture sunlight and increase body temperature to lay eggs (James 1988). In this study, the south-facing leaf litter was exposed to sunlight during the daytime, hence the preference of the kudzu bugs was justified. As observed by Kuzin et al. (1980), Telenomus species seek overwintering hosts in woodland, orchard and forest environment probably to overwinter in their vicinity and parasitize their eggs as soon as they oviposit in spring. 1 specimen each of Scelionidae was found in leaf litter and bark in this study (Table 3).
Objective Two
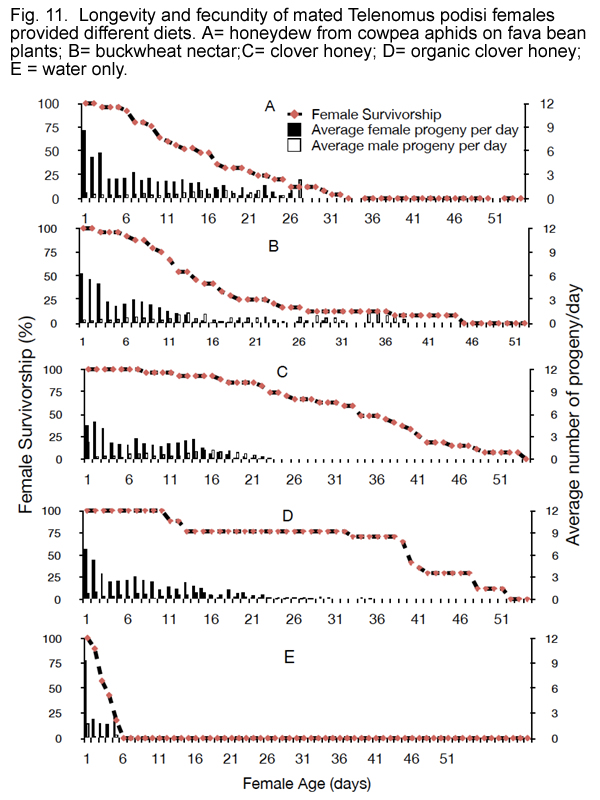
There was a significant effect of different diets on the longevity of T. podisi (F-value = 25.53, df = 21, 99, P < 0.0001) (Fig. 11). Mean longevity of T. podisi fed on clover honey (either conventional or organic) was approximately 2-fold higher than those fed aphid honeydew and buckwheat nectar, and 11-fold higher than starved wasps (P < 0.0001). Interestingly, mean longevity of T. podisi fed on aphid honeydew and buckwheat nectar was not statistically different (P = 0.668; Table 4).
Fecundity of T. podisi was also significantly affected by the different diets (F-value = 5.93, df = 21, 99, P < 0.0001). There was no statistical difference between mean fecundity of T. podisi fed on aphid honeydew, buckwheat nectar, conventional clover honey and organic clover honey (P ? 0.05; Table 4) (Fig. 11). Fecundity of wasps fed on these diets was 3 to 4-fold higher than that of starved wasps (P < 0.0001).
There was no significant effect of different diets on the progeny sex-ratio of T. podisi (F-value = 1.54, df = 21, 95, P = 0.0839) (Table 4). There was a strong day effect over time (Wald Chi-square = 35.14, P < 0.0001) (Fig. 11), with estimate = -0.1226 (±0.02). Wasps fed the various diets produced female progeny for an average of 26.8 ± 2.2 days and male progeny for 30.8 ± 6.9 days with the average sex-ratio on the first day of oviposition being 0.9 ± 0.1 and that on the last day being 0.4 ± 0.1.
These parasitoids continued to kill host eggs through attempted oviposition even after they had stopped producing progeny, although fewer host eggs were killed (F = 38.08, df = 1, 120, P < 0.0001). When producing progeny, the wasps killed 98.9% of the 8785 host eggs presented to them while wasps no longer producing progeny killed 87.9% of the 6330 eggs presented to them.
Figure 11 shows that aphid honeydew is as beneficial as nectar or honey in terms of enhancing the fecundity and longevity of Telenomus podisi. This is a deviation from other studies dealing with honeydew and parasitoids which concluded that honeydew is typically an inferior food source compared with artificial sugar-rich foods and some floral nectars because it contains sugars that do not readily elicit a feeding response or are nutritionally less suitable (Lee et al., 2004; Wäckers, 2001). This may be explained by our presenting honeydew in situ (i.e. on living leaves with aphids present) as opposed to collected directly from aphids. It was found by Fuchsberg et. al. (2007) that effect of honeydew from corn leaf aphid, Rhopalosiphum maidis (Homoptera: Aphididae) on Trichogramma ostriniae (Hymenoptera: Trichogrammatidae) allowed increase in survival and parasitism rate when compared to only corn leaf or nothing provided in experimental cages. The aphids were offered in situ in this study as well. Additional studies are currently underway to explore the possibility of phyllosphere yeasts enhancing nutritional value of honeydew.
References:
Fuchsberg, J. R.; Yong, T.; Losey, J. E.; Carter, M. E. and Hoffmann, M. P. 2007. Evaluation of corn leaf aphid (Rhopalosiphum maidis; Homoptera: Aphididae) honeydew as a food source for the egg parasitoid Trichogramma ostriniae (Hymenoptera: Trichogrammatidae). Biological Control 40: 230–236.
James, D. G. 1988. Fecundity, longevity and overwintering of Trissolcus biproruli Girault (Hymenoptera: Scelionidae) a parasitoid of Biprorulus bibax Breddin (Hemiptera: Pentatomidae). Australian Journal of Entomology 27 (4): 297 – 301.
Kuzin, A. A.; Kudryavtsev, V., S. and Ponurovskii, A. Y. 1980. Noxious bugs on wheat and their insect enemies. Zashchita Rastenii (10): 24-25.
Lee, J.C.; Heimpel, G.E. and Leibee, G.L. 2004. Comparing floral nectar and aphid honeydew diets on the longevity and nutrient levels of a parasitoid wasp. Entomol. Exp. Appl. 111, 189–199.
Wäckers, F. 2001. A comparison of nectar and honeydew sugars with respect to their utilization by the hymopteran parasitoid Cotesia glomerata. J. Insect Physiol. 47: 1077–1084.
Witting-Bissinger, B. E.; Orr, D. B. and Linker, H. M. 2008. Effects of floral resources on fitness of the parasitoids Trichogramma exiguum (Hymenoptera: Trichogrammatidae) and Cotesia congregata (Hymenoptera: Braconidae). Biological Control 47: 180–186.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Educational & Outreach Activities
Participation summary:
Lahiri, S. Potential for conservation biological control of stinkbugs in North Carolina. Ph.D. Dissertation, N.C. State University, Raleigh, NC. (In progress).
Lahiri, S., D. Orr, C. Sorenson, A.C. Cohen and Y.J. Cardoza. 2013. Comparative value of various carbohydrate sources for the stink bug egg parasitoid Telenomus podisi (Hymenoptera: Scelionidae). Entomological Society of America annual meeting, Nov. 10-13, 2013, Austin, Texas. (10 minute presentation).
Lahiri, S., D. Orr, C. Sorenson, A.C. Cohen and Y.J. Cardoza. 2013. Potential overwintering refuge sites for kudzu bugs (Hemiptera: Plataspidae) and parasitoids. Entomological Society of America annual meeting, Nov. 10-13, 2013, Austin, Texas. (poster presentation).
Lahiri, S. 2012. Conservation biological control of stinkbugs. 20 minute presentation as part of: CEFS Seasons of Sustainable Agriculture Series Workshop “Habitat for Multiple Ecological Services”, Center for Environmental Farming Systems, Goldsboro, May 23, 2012. (37 attendees; half-day workshop)
Lahiri, S. 2012. Potential for Conservation Biological Control of Stink Bugs in North Carolina". North Carolina State University Entomology Graduate Student Symposium, January 26-27, 2012.
Lahiri, S. 2011. Conservation biological control of stinkbugs. 20 minute presentation as part of: CEFS Seasons of Sustainable Agriculture Series Workshop “Farmscaping For Pest and Wildlife Management”, Center for Environmental Farming Systems, Goldsboro, May 25, 2011. (34 attendees; half-day workshop)
Presented information on conservation biological control of stinkbugs as part of a booth on biological control and IPM at BugFest, Museum of Natural Sciences, Raleigh, Sept. 17, 2011, Sept. 15, 2012, Sept. 21, 2013.
Project Outcomes
The results of this study indicate that in the field, T. podisi may be able to benefit equally from nectar as well as honeydew which is ubiquitously present in agro-ecosystems. Field and lab overwintering studies to date suggest the importance of woodland field borders with leaf litter from hardwood trees as a refuge area during winter for scelionid parasitoids and kudzu bugs. This study, when complete should help to guide future conservation biological control efforts with both native and imported scelionid egg parasitoids of stinkbugs and their relatives (e.g. kudzu bugs).
Economic Analysis
N/A
Farmer Adoption
This project was intended to improve understanding of insect life histories on farms with a long-term goal of improving biological control of stink bugs and their relatives. Immediate adoption of specific practices was not intended, but it was intended that this work will help development of specific practices that will be adopted by farmers in the future.
Areas needing additional study
An examination should be conducted of microbial activity on leaf surfaces (phyllosphere) and possible enhancement of honeydew quality for parasitoids.