Final Report for GS11-106
Project Information
A two-year trial to study the efficacy of providing artificial nest boxes to increase bumble bee abundances on farm sites was implemented in 2012 and 2013. None of the 300 nest boxes deployed over two years was occupied by bumble bees, regardless of insulation material, entrance type or orientation. The phenology of bee-visited crop blooms was tracked and compared to the phenology of bumble bee species in the area. Bumble bees active in June through August visited more crops than those active earlier in the season.
Introduction
With recent honey bee declines impacting managed pollination services, the vulnerability of American agriculture under the dominance of the honey bee has become increasingly apparent in recent years. Many American farms utilize managed honey bees for crop pollination services, yet dependence upon a single, non-native organism for such a crucial service is an unsustainable approach. Bumble bees are native pollinators that visit a wide variety of plant species and can pollinate many field crops such as berries, melons, cucumbers, etc (reviewed in Klein et al. 2007). Wild populations of native bumble bees freely provide indispensable pollination services which have been shown to surpass those of honey bees in many crops (e.g. watermelon, Winfree et al. 2008). However, in some agricultural systems, their pollination efficiency seems to be limited by their population size (e.g. pak choi, Rader et al. 2009). In order to be effective crop pollinators, bumble bees must not only be abundant enough to provide ample pollination services, but their seasonal cycles must also coincide with crop bloom periods. Bumble bee species exhibit a seasonal colony cycle, with some species appearing early in spring and declining by mid-summer, and others having the opposite cycle (Colla and Dumesh 2010). Clearly data on the seasonal abundances of regional bumble bee species could aid farmers seeking to improve native pollination services. One of the grassroots recommendations for enhancing bumble bee populations pollination services is adding artificial nest boxes to increase local populations (e.g. Mader et al. 2010). Reported occupancy rates of nest boxes vary widely, ranging from 0% to 67% occupancy, but the real utility of this method for attracting and maintaining bumble bees has not often been tested in the USA southeastern United States (Holm 1966; Donovan and Wier 1978; Delaplane and Mayer 2000). Our study aimed to develop recommendations for providing bumble bee nest boxes (hereafter, “boxes”) to increase native pollinator abundance and provide information on the timing of bumble bee abundances in relation to crop bloom to aid farmers in their ability to capitalize on native pollinators.
1. Evaluate the utility bumble bee nest boxes for increasing the prevalence of bumble bees on small, multi-crop farms 2. Determine if providing additional bumble bee nesting sites via artificial nesting boxes changes the species diversity and relative abundance of bumble bees 3. Determine the seasonal abundance of different bumble bee species as it relates to crop bloom times 4. Develop a set of recommendations for bumble bee nest boxes for farmers in Arkansas
Cooperators
Research
A local farmer in Northwest Arkansas agreed to be a collaborator on this project. This farmer has approximately 20 acres of crops and grows fruit and vegetables for local markets. His operation is diverse, incorporating row, greenhouse and high tunnel production of tomatoes, peppers, strawberries, squash, pumpkins and a variety of other fruits and vegetables. Preliminary trials were conducted on this farm and the University of Arkansas Experimental farm in 2012. In 2013, an inundative trial was conducted on the local collaborator’s farm.
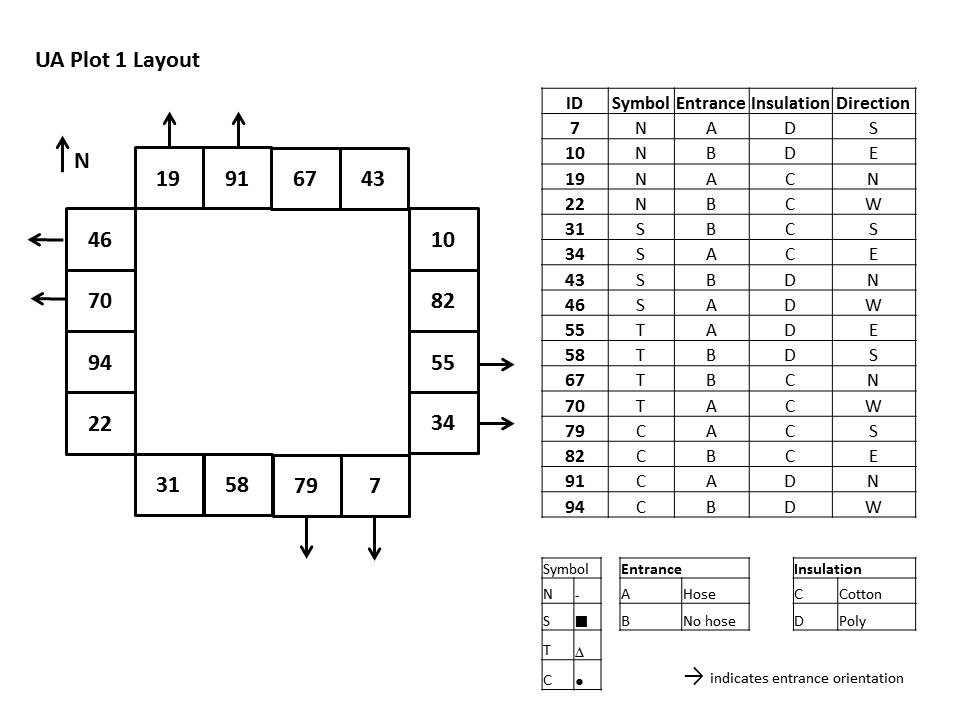
The wooden, two-chamber design of Welland (1995) was chosen and determined to be an efficient artificial nest design for the boxes. Feet were also added to help improve airflow and control moisture (Hobbs 1967). Two types of insulation materials, bamboo-rayon-poly fiber (Fig. 1a) and recycled cotton insulation were implemented (Fig. 1b). Because the literature on natural nests mentions that many bumble bee nests have long entrance tunnels (Plath 1922; Donovan and Wier 1978), two entrance designs were implemented. One treatment included boxes with entrance tunnels constructed from buried hoses (Fig. 1c) and the other treatment had a plain entrance hole on the face of the box (Fig .1d). Because the boxes were deployed in groups, it was decided that orientation may be an issue for bees upon occupancy. Four symbols were chosen to help differentiate the boxes from one another: 1) circle (C), 2) square (S), 3) triangle (T) and 4) no symbol (N), adding a blocking factor to the design. The primary factors were 1) Entrance type: hose (A) or hole (B), 2) Insulation type: cotton (C) or bamboo-poly (D), and 3) Cardinal direction: North, South, East or West. Thus each replicate consisted of 16 nest boxes, one of each available combination of factors (see Fig 2 for an example of a replicate layout incorporating all factors). In the preliminary trials of 2012, three replicates (48 boxes) were installed in different locations > 50 m apart on each farm in March. In 2013, 12 replicates (192 boxes) were installed on the collaborator’s farm in February. An additional 12 boxes were installed containing nest material constructed to mimic a rodent’s nest (Mjelde, 2013). Occupancy was monitored twice a month until the first frost, at which point all boxes were removed.
Bumble bee diversity and abundance were recorded at the collaborator’s farm with standardized surveys between late March and mid-October in both 2012 and 2013. Surveys were conducted by walking at a slow, even pace through the patches of flowers, both crop blooms and field margins, present at the site. Over a 30-min period all bumble bees observed within approximately 10 m of the observer were netted with an aerial net. The number of individuals, their species, caste and floral hosts were recorded. Most of these individuals were subsequently released, although some were retained as vouchers. Others that were too worn to be reliably identified in the field were also retained for subsequent identification to species using the keys of Mitchell (1962) and Chandler and McCoy, Jr. (1965). Because they are crucial to local population growth throughout a season, queens were not retained. Crops that were in bloom on site were recorded during each site visit. Crop bloom data from another farm site nearby (9.1 km) were added as well to provide additional data on bloom phenology in the area.
Bumble bee diversity on the collaborator’s farm was assessed through comparison with diversity data collected with the same methodology as a part of a concurrent, unpublished study conducted by the graduate student researcher in the Northwest Arkansas region. The Shannon-Wiener diversity index (H’) was calculated (Hines & Hendrix 2005) to allow comparisons of diversity between the farm in this study and non-farm sites (n = 11) to gauge on-farm diversity. These data were also used to construct a local bumble bee phenology to determine which species were present during crop bloom. Bumble bee floral visitations to crop and non-crop plants on the farm were visualized with a bipartite network graph (Dormann, et al. 2008) in R (R Core Team, R Foundation for Statistical Computing, Vienna, Austria).
- Figure 1. Artificial nest box design and treatments employed in this study. a) bamboo-rayon poly insulation; b) recycled cotton insulation; c) installed box with a buried hose entrance; d) installed box with a simple entrance hole.
- Figure 2. An example of the layout of boxes used within each replicate and treatment factors employed. Each treatment factor combination was used once per replicate.
{kind=link}
{kind=link}
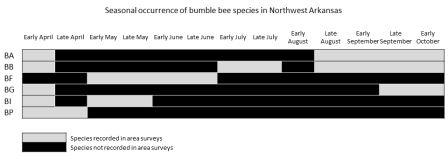
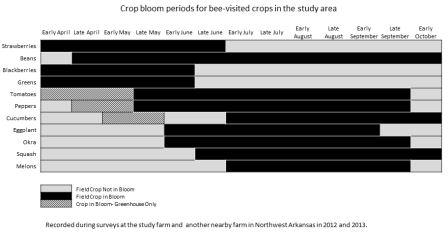
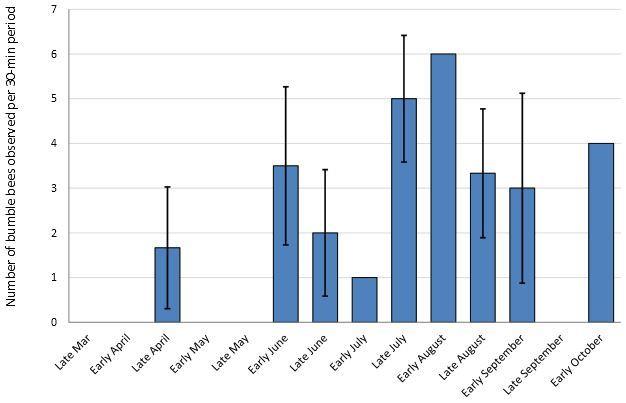
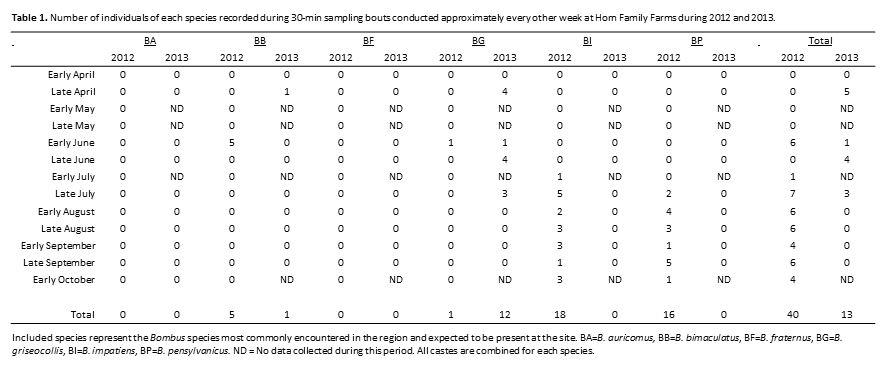
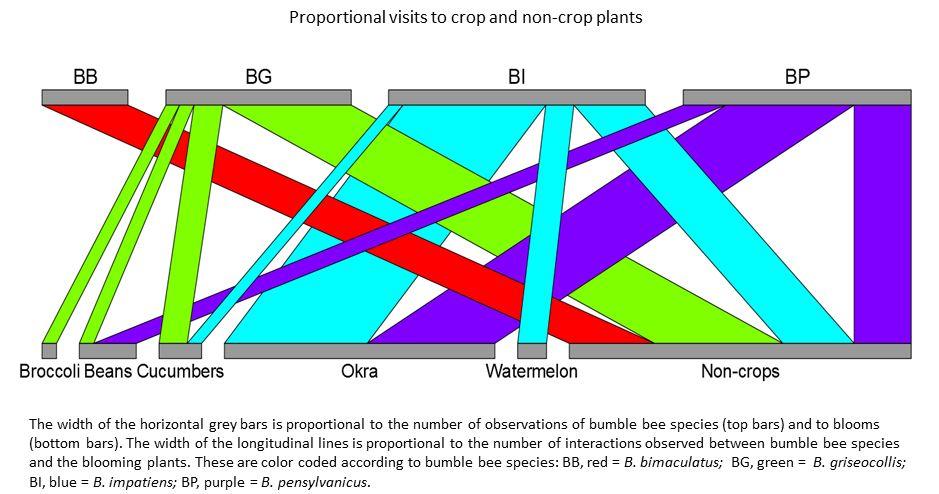
No artificial nests were inhabited by bumble bees in either 2012 or 2013, regardless of treatment. Other organisms did utilize the nests including Polistes wasps (Fig. 3a), Megachilidae bees (Fig. 3b), Crematogaster (Fig. 3c) and other ants and mice (Fig. 3d). A single natural bumble bee nest was observed on the site in 2013. The B. griseocollis colony was located in an underground rodent’s nest about 1 m away from one of the artificial nest boxes. Twenty-four surveys of bumble bee presence on the farm were conducted over the two-year study period. During each 30-min sampling bout, 0 - 7 individual bumble bees were observed, with an average 2.30 ± 0.54 bees observed per bout. Overall bumble bee abundance per visit peaked in late summer, with the most bees observed per visit in early August (Fig. 4). Of the six bumble bee species in the area (B. auricomus, B. bimaculatus, B. fraternus, B. griseocollis, B. impatiens and B. pensylvanicus), only four occurred at the site (Table 1). The two species that were not recorded at the farm site, B. auricomus and B. fraternus, were also the least common species recorded at non-farm sites throughout the region in 2012 and 2013 (5.1% and 3.7% of the total observations in the region, unpublished data). Even though only four species were recorded at the farm site, the Shannon-Weiner diversity index for the farm site equaled that of all non-farm sites combined (H’ = 1.90). Bumble bees were observed in the region from April through October, although not all species were present during all periods (Fig. 5). The only species present in early April was the less common B. fraternus, which was not observed on the farm site. Crop bloom periods were recorded for eleven crops common on the two farm sites and visited by bees (both bumble bees and honey bees, data not shown). Bee-visited crops were also blooming from April through October (Fig. 6). Bumble bees were not observed on all crops, however. Bumble bees were observed on five crops, broccoli, beans, cucumbers, okra and watermelon, as well as on non-crop plants present at the site (Fig. 7). Bombus bimaculatus was not observed on any of the blooming crop species, and B. griseocollis was observed on non-crop blooms more often than on crop blooms (67% and 33% of visits, respectively). Both B. impatiens and B. pensylvanicus were recorded on crop blooms more frequently (72% and 75%, respectively) than non-crop blooms. Bombus griseocollis was the only bumble bee species recorded visiting broccoli blooms, and B. impatiens was the only species recorded on watermelon blooms.
None of the artificial nest boxes in this study were occupied by bumble bees. Although artificial nests are often recommended as a means of increasing local bumble bee populations, this tactic was ineffective in our farm site. The reason for the lack of occupancy is unknown, but a number of factors likely influence nest site choices of bumble bee queens. In studies in other countries, artificial nests of many designs have had varying rates of occupancy with no consensus on the optimal design (e.g. 0-7% in Britain, Lye, et al. 2011; 24.7% in New Zealand, Donovan & Weir 1978; at least 20% in Canada, Hobbs 1967). Large, formal studies on artificial nest boxes in the United States are scant, but thus far, occupancy rates in regions outside of the southeast are just as dismal. A trial conducted in Pennsylvania with multiple locations, designs and placements had an occupancy rate of 1.6% (Bogdash 2009). In Nebraska, 400 boxes designed and installed by high school science teams had zero occupancy in state-wide trials (Golick & Ellis 2003). Our study provides evidence that artificial nest boxes may similarly unsuitable in the southeast, although it is possible that the basic design that we chose was unsuitable for local species. More information on natural nest-site selection by queens could prove useful in designing more attractive artificial nests. Even with an ideal design, however, artificial nests can only be effective in increasing bumble bee populations if nesting is limited at a particular site. It is possible that natural nest sites were sufficient for the local bumble bee population at our farm, rendering our boxes less attractive than natural nesting sites already available. Other factors may also play a role in governing bumble bee abundances on farms, such as the availability of floral resources throughout the long season of bumble bee activity. Bumble bees are present from April through October in the area. On multiple crop farms, such as the one implemented in this study, crop plants are in bloom throughout the entire active season of bumble bee species in the area. This should help provide food resources to maintain bumble bee populations better than farms that focus on a single crop with a limited bloom period. Bumble bee visitation to crop blooms was low overall, but visitations increased for crops blooming between June and August when the later season species B. impatiens and B. pensylvanicus were most active. Bombus bimaculatus was an early season species active between late April and late June and was not recorded foraging on crop blooms. The earliest recorded species in the region, B. fraternus, was not present on the study farm. Although these data are preliminary, they do suggest that early season crop blooms may be less likely to receive pollination by bumble bees in the area.
- Figure 3. Examples of non-bumble bee occupants of artificial nest boxes. a) a newly constructed Polistes wasp nest; b) nests of Megachilid bees; c) Crematogaster ant colony; d) a mouse nest.
- Figure 5. The seasonal occurrence of bumble bee species in Northwest Arkansas in 2012 and 2013. All castes are combined. BA = B. auricomus; BB = B. bimaculatus; BF = B. fraternus; BG = B. griseocollis; BI = B. impatiens and BP = B. pensylvanicus.
- Figure 6. Blooming periods for some bee-visited crop plants in the area. Note that although greens do not require pollination for marketable product, their blooms can be a useful food resource for bumble bees if allowed to remain in the field. All other crops listed here are assumed to require pollination to set marketable product.
- Figure 4. The number of bumble bees observed during a 30-min sampling bout over each period throughout the season. All farm site visits during each period in 2012 and 2013 (n = 1 – 3 per period) were averaged. Periods that had more than one visit have standard errors shown as bars.
- Number of individuals of each species recorded during 30-min sampling bouts conducted approximately every other week at Horn Family Farms during 2012 and 2013.
- Figure 7. Visitation of bumble bees to crop and non-crop plants on the study farm in 2012 and 2013. Note that although broccoli is harvested prior to blooming, blooms can be a valuable food resource for bumble bees. All other crops are assumed to require pollination to set harvestable product.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Educational & Outreach Activities
Participation summary:
2011. Creating habitats for native pollinators. 1.5-hour presentation to the Fayetteville Community Wildlife Habitat Project, Fayetteville, AR. 2012. Good and bad bugs of the garden. 1-hour workshop presentation to the Dig In! Food and Farming Festival, Fayetteville, AR; with Soo-Hoon Sam Kim. 2012. Bumble bee domiciles. 15-minute workshop presentation as part of the Arkansas Pollinator Conservation Short Course Hosted by the Xerces Society for Insect Conservation, Booneville, AR; with many others. 2012. Native bees. 1-hour presentation to the Garfield Garden Club Annual Meeting, Fayetteville, AR.
Data from this study will be combined with data from additional studies to prepare a manuscript describing the seasonal abundance of bumble bee species and bloom phenology in the region.
Project Outcomes
The main outcome from this project is that efforts to increase bumble bee populations on farms for pollination services should not rely on providing artificial nest boxes. These do not seem effective in the region. Instead, efforts should focus on other factors known to influence bumble bee abundances, such as providing floral resources throughout the entire period of bumble bee activity. Multiple-crop farms such as the one employed in this study seem to provide blooms throughout this period. In northwest Arkansas, bumble bees are active from April through October and should be capable of lending pollination services to farmers in the region over much of the growing season, although increasing abundances on farms is desirable In this particular system, bumble bee visits to crop blooms were greatest from June through August and less in the early season from April to June. This indicates that efforts to increase bumble bee pollination services may be best directed at early season species such as B. bimaculatus.
Areas needing additional study
Although this study has shown that nest boxes are an ineffective means of increasing bumble presence on farms, it was unable to determine why. Studies that address whether or not nest sites are limiting in agricultural systems should prove useful in future artificial nest box endeavors. It is also possible that providing a more suitable artificial nest may help boost bumble bee numbers on farms. The factors that influence nest site selection in bumble bee queens are largely unknown. Determining what makes a nest site attractive to site-seeking queens may yield better results in future artificial nest box trials. Additional studies are required in different regions of the United States to determine if nesting boxes are more successful in these areas.
Bogdash, S. 2009. NE-SARE Report ONE08-079. Available online at https://projects.sare.org. Chandler, L. and C. E. McCoy Jr. 1965. Arkansas Academy of Science Proceedings 19: 46-53. Colla, S. R., and S. Dumesh. 2010. Journal of the Entomological Society of Ontario 141: 39-68. Delaplane, K. S. and D. F. Mayer [eds.]. 2000. Crop Pollination by Bees. CABI Publishing, New York, NY. Donovan, B. J. and S. S. Wier. 1978. Journal of Agricultural Research 21: 733-756. Dormann, C.F., B. Gruber and J. Fruend. 2008. R News 8(2): 8-11. Golick, D. A. and M. D. Ellis. 2003. American Entomologist 49(2): 74-79. Hines, H. M. and S. D. Hendrix. 2005. Environmental Entomology 34(6): 1477-1484. Hobbs, G. A. 1967. Canadian Entomologist 99(9): 943-951. Holm, S. N. 1966. Annual Review of Entomology 11: 155-182. Klein, A. M., et al. 2007. Proceedings of the Royal Society B-Biological Sciences 274: 1-35, Appendix 2. Lye, G. C., K. J. Park, J. M. Holland, and D. Goulson. 2011. Journal for Nature Conservation 19: 154-160. Mader, E., M., et al. 2010.ATTRA-National Sustainable Agriculture Information Service. Available online at https://attra.ncat.org/attra-pub/summaries/summary.php?pub=75. Mitchell, T. B. 1962. Technical Bulletin 152. North Carolina Agricultural Experiment Station, pp. 557. Mjelde, A. 2013. Fagbladet Oko Logisk Landbruk 1: 36-37. Plath, O. E. 1922. Psyche 29(5-6): 189-202. Rader, R., et al. 2009. Journal of Applied Ecology 46: 1080-1087. Welland, R. 1995. B.C. Fruit Testers Association 8(1): 13-14. Winfree, R., et al. 2008. Journal of Applied Ecology 45: 793-802.