Final report for GS16-154
Project Information
As the primary source of organic compounds in terrestrial ecosystems, plants have developed a striking diversity of morphological and biochemical mechanisms to reduce the impact of herbivory. Plant defenses strongly select for resistance in herbivorous insects, leading to a high degree of specialization. Subsequently, resistant herbivores utilize host-specific ratios of emitted volatile organic compounds to discriminate between suitable and unsuitable potential hosts. In this study we attempted to elucidate the factors involved in long-range oviposition site selection in the peachtree borer (Synanthedon exitiosa) and lesser peachtree borer (Synanthedon pictipes). In doing so, we hope to facilitate the development of female-oriented monitoring and mass trapping strategies based on the chemical ecology of these economically significant pests. Accomplishing this goal required us to:
- Identify plastic traits associated with host quality;
- Determine what long-range chemical cues are associated with these traits; and
- Assess the degree to which these cues are suitable for attracting sesiids in the field.
The literature suggests that both sesiid peach pests lay more eggs at sites where bark has been damage (Cottrell et al., 2008; Derksen et al., 2006). As wounding is associated with changes in local and systemic volatile emissions, we determined that this trait was suitable. Thereafter we:
- Profiled the constitutive volatile emissions from bark/phloem samples of peach (Prunuspersica), black cherry (Prunus serotina), and apple (Malus pumila) using static HS-SPME GC-MS;
- Documented shifts in the emission of various defensive compounds twenty-four hours after mechanical wounding; and
- Determined whether synthetic blends, mimicking the natural volatile emissions of wounded bark effectively attracted PTB or LPTB females.
We report that mechanical damage affected the emission rate of terpenes, terpene-derived compounds, and benzenoids. Notably, benzaldehyde, p-cymene, camphene, and linalool emission underwent similar shifts in both host species examined. This suggests that one or more of these compounds conveys information about the physical impediments likely to be encountered by larvae colonizing a tree.
Samples collected from cultivated peach trees in the defunct Collins Round Mountain Orchard in Conway, AR (field site) failed to show differences in the rate of volatile emission before and after mechanical wounding. The constitutive profile of these trees also showed dramatically reduced benzaldehyde emission alongside increased diversity and concentration of the sesquiterpenes guaiene, cadinene, caryophyllene, humulene, and farnesene compared to wild peach trees. We attribute this to the fact that healthy individuals were sampled from among trees facing microbial infestation.
Samples collected from the non-host species, apple, emitted two monoterpenes, camphene and alpha-pinene, at higher rates. However, the ratio at which these compounds were emitted strongly favored the former, whereas in Prunus species the compounds were emitted at equal rates. Apple was the only species to emit the sesquiterpene alpha-bergmotene and lacked the sesquiterpene diversity observed in the Prunus species.
Although pheromone trapping was highly successful, neither the kairomone blend based on cultivated peach samples or the blend based on wild peach were suitable for attracting female sesiids. In fact, the distribution of early instar larvae observed in July and August of 2017 may suggest the opposite. The distribution of early instar PTB larvae was mapped to indirectly confirm female activity in the area. In areas where kairmone lures were present, the percent of trees infested was reduced by half. Although the current study was not designed to address the question of deterrence, it may represent a worthwhile avenue for future research.

This project examines the interaction of adult female PTBs and the complex mixture of volatile organic compounds emitted from optimal and suboptimal oviposition sites, with the expectation that such information will inform the development of female-oriented PTB lures. Accomplishing this goal requires the integration of analyses from the realms of metabolomics, animal behavior, and electrophysiology in the laboratory to develop attractive lures, and a subsequent field test of lure efficacy.
With this in mind our primary objectives were initially:
- Identify the volatiles that are likely involved in host selection in the PTB.
- Determine the bioactivity of these volatiles.
- Evaluate the extent to which these volatiles are behaviorally relevant in the lab.
- Test the efficacy of these compounds as lures in the field.
Cooperators
Research
Plants and Sampling
Samples of cultivated peach and apple were collected from the defunct Collins Round Mountain Orchard in Conway, Arkansas, 18-24 months after operations had ceased. Wild peach and black cherry were sampled from Emerald Park in North Little Rock, Arkansas. Cultivated trees were selected from within a 1000 m2 area and of similar condition. Two strips of bark/phloem tissue, measuring approximately 2.5 by 1.25 cm and weighing approximately 0.5 grams each, were cut from opposing sides of each of six trees and snap frozen in liquid nitrogen. The wounds were subsequently sealed with parafilm and approximately twenty-four hours after the initial sampling, strips of bark/phloem adjacent to wound sites were collected as previously described. Upon returning to the lab, samples were transferred to 10 ml headspace vials and weighed, before being stored at -85°C until extraction.
HS-SPME GC-MS Analysis
The fiber utilized in this study was composed of divinylbenzene (DVB) and carboxen (CAR), with a coating of polydimethylsiloxane (PDMS), which is the recommended fiber for low-medium molecular weight VOCs and flavor components. Samples were allowed to warm to room temperature, at which time solid-phase microextrations of the headspace above the bark/phloem samples were conducted manually. The SPME fiber was inserted through a rubber septum and exposed from a needle depth of 0.5 cm for 30 minutes without agitation. Subsequently, volatiles were thermally desorbed over the course of 2 minutes at 250 C in the split-/splitless inlet (in splitless mode) of an Agilent 6890 gas chromatograph coupled to an Agilent 5975 mass spectrometer. The needle depth during desorption was 4 cm.
Oven parameters: The initial temperature was held at 30 C for 5 minutes to recondense the analytes in the column head before being increased to 150 C at 10 C/minute. The oven was further ramped at 25 C/minute to 250C, where it was held for 2 minutes.
MSD parameters: The mass spectrometer began scanning from 40 to 350 amu after a solvent delay of 5 minutes. Compounds were ionized via electron impact at 70 eV.
Electoantennography
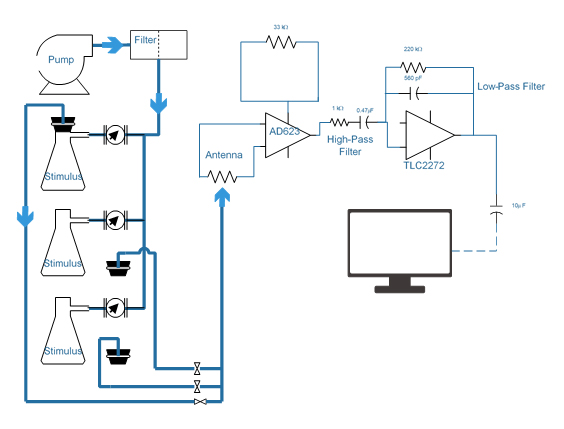
A device based on schematics from Bjostad and Roelofs (1980) was constructed using materials sourced through Backyard Brains (Ann Arbor, MI). The device amplified the antennal signal 696-fold and displayed the trace on a laptop.
Male peachtree borers were collected by net on 25 June 2017 at the location of a pheromone lure. These animals were stored in a glass jar at approximately 78°F until use. Before analysis moths were chilled to facilitate handling and coaxed into micropipette tips, becoming trapped with their antennae and heads exposed. Each moth was decapitated and the tip of one antenna was inserted into a glass capillary electrode, while another electrode was inserted through the eye. Conductive gel (Spectra 360) ensured contact between the EAG circuit and antenna.
Each antenna was allowed to acclimate to the stream of charcoal filtered, humidified air, as the chemicals were prepared. Observed voltage deflections during this time period suggested that the device’s sensitivity was adequate. 50 mg of each compound was loaded into rubber septa and housed individually in Büchner flasks. Air continuously flowed into the flasks through the hose barb, but the stream from each flask was only introduced to the main airstream when the flask was briefly sealed. This ensured that the antenna received similar concentrations of each compound.
Compounds of interest were introduced every forty-five seconds, when the appropriate flask was sealed for fifteen seconds. It was expected that a strong initial deflection, followed by lower magnitude depolarizations as the antennal neurons became habituated to the stimulus, would be observed over the course of the exposure period.
After we observed no response to benzaldehyde, which Derksen et al. (2007) reported induced a response in male PTBs, sex pheromone was tested as a positive control.
Field Trapping
After chromatographic analysis was performed, a mass trapping experiment was conducted at the defunct orchard. Each of thirty 225 m2 plots were baited with either peachtree borer pheromone (Great Lakes IPM), 5 mm rubber septa loaded with clean dichloromethane (DCM), or septa loaded with 100, 200, or 300 mg of a blend of VOCs identified in the orchard bark samples. Lures persisted in the orchard for seven weeks, between 21 June 2017 and 9 August 2017, with traps being replaced as necessary.
When the volatile profile of cultivated peach bark was observed to be atypical, an additional eighteen traps were baited with pheromone, DCM, or 100 mg of a second potential kairomone blend (based on wild peach emissions). The second round of trapping began 20 July 2017 and continued for three weeks until 10 August 2017.

Assessment of Host Quality
Pheromone traps were set in two locations, the defunct Collins Round Mountain Orchard in Conway, AR and suburban Little Rock, AR, in mid-May 2017. The emergence patterns and size of male PTBs were documented between 20 May and 14 August 2017. Emergence patterns were documented as weekly capture rates and totals, which were compared across sites based on degree-days (Johnson and Mayes, 1983). Body length and wingspan of mounted specimens were measured with metric dial calipers. Size comparisons were based on the measurements of forty-five individuals from each site that were collected during the same eclosion event (i.e., a large number of adults appearing in a short time after a lull).
Assessment of Larval Distribution
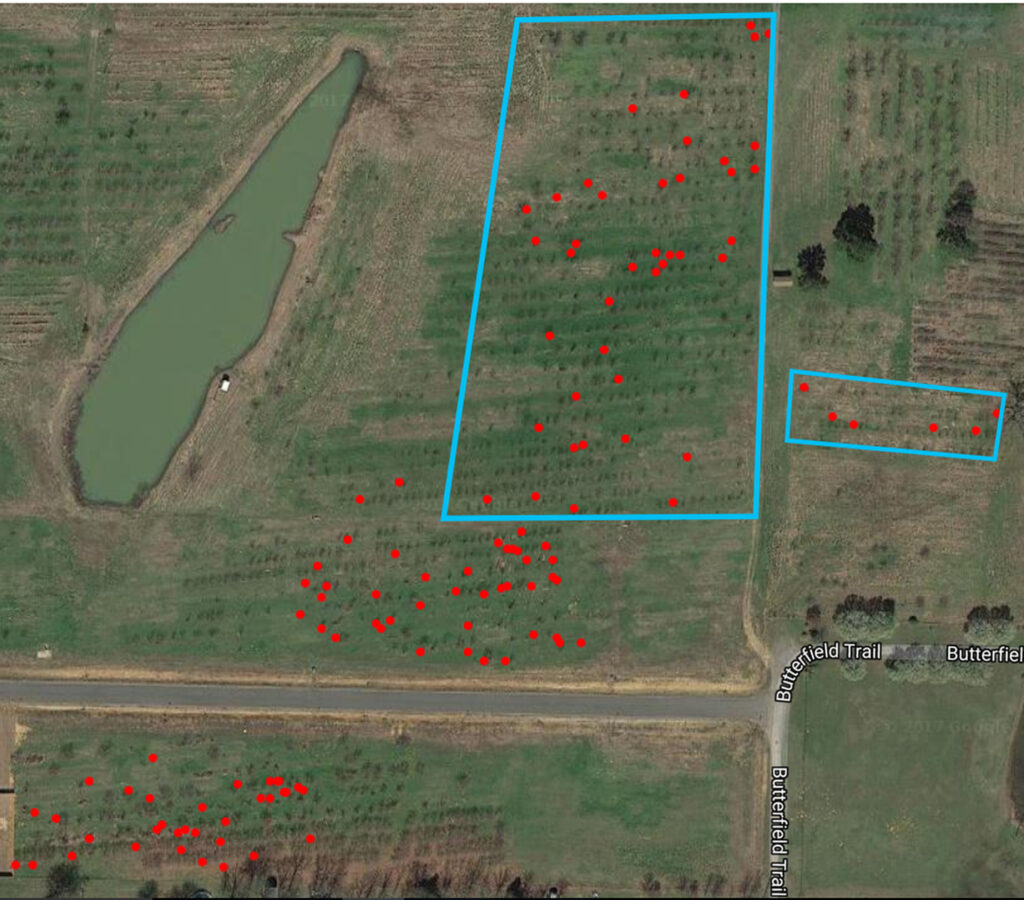
Over the course of three weeks, between 30 July and 13 August 2017, trees in three large plots were examined for the presence of PTB larvae. When larvae were discovered, they were classified as either ‘Early-’, ‘Middle-’, or ‘Late-Instar’ based on size, and flagged. On 12-13 August 2017 all unflagged trees were re-examined and the coordinates of each juvenile (early- or middle-instar) larva was documented using a handheld Garmin GPS 72 and plotted using google maps. Additionally, the percentage of infested trees was calculated for areas with and without kairomone lures.
Data Processing and Analysis:
Chromatography
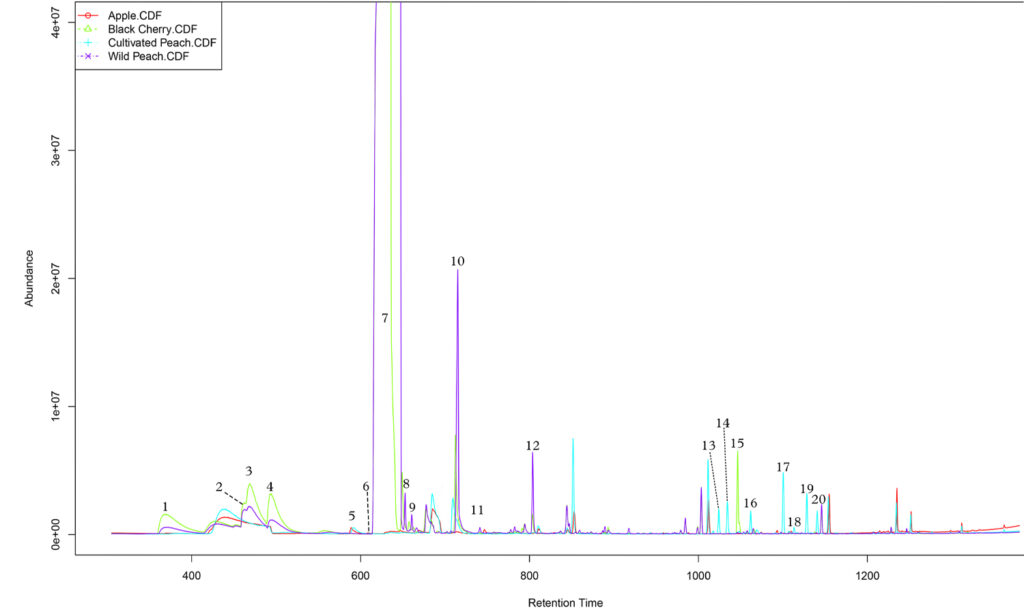
Preliminary analysis of chromatograms was performed via the data acquisition software. A sub-library of tentatively identified compounds, those returning a greater than 85% library match, was generated utilizing the Wiley 7th Edition database. Subsequent data processing and deconvolution (Stein, 1999) was performed in AMDIS (Automated Mass Spectral Deconvolution and Identification System) with target compounds defined by the previously generated sub-library.
The data generated via AMDIS batch processing were analyzed in R using Metab (Aggio, 2011) to generate matrices of peak data that were normalized by sample mass. For improved clarity, data frames comparing each treatment were generated. The resultant data frames were log-transformed and principle component analyses were conducted for each pair of treatments (i.e. cherry vs. peach after wounding or peach before vs. peach after wounding). PCA loadings were then examined to assess the degree to which each peak contributed to the overall variance. Those compounds that contributed strongly to variance between host and non-host species, but weakly to the variance between potential hosts, were considered promising candidates in facilitating host selection in peachtree and lesser peachtree borers. Subsequently, Student t-tests were used to assess differences in the emission of each metabolite. Having seen clear discrimination in the PCA, and being somewhat worried about our limited sample size (restricted by the availability of apple and wild prunus species), we elected to set alpha equal to 0.1 for these tests to reduce the likelihood of a type II error.
After statistical analysis, a standard blend to fatty acid-methyl esters (C8-C24 from Sigma-Aldrich) was used to calculate the Kovats retention index (RI) of each peak. The RI of each tentatively identified compound was compared to published values from the NIST webbook (NIST Mass Spec Data Center). The identity of each compound was considered to be confirmed if the observed RI was within 1% of the published value.
Trapping
Field data were examined with regard to both total captures and weekly capture rates. ANOVA was used to compare these measures among the negative control (DCM), positive control (pheromone), and experimental (kairomone) groups.
Host Quality
It was determined that length data were independent, normally distributed, and possessed equal variances, allowing for the use of a Student t-test. Assumptions of parametric analysis were not met for wingspan measurements, resulting in the use of a Wilcoxon Rank-Sum Test for comparison.
Larval Distribution
The extent of infestation was compared across three plots (an area where trapping occurred, an adjacent area, and a more distant area) using a Chi-Square test for equality of proportion.
A variety of secondary metabolites including green leaf volatiles (GLVs), terpenes, and benzenoids were detected in the headspace above peach, black cherry, and apple bark/phloem samples. However, the synthetic kairomone blends derived from these observations proved ineffective for monitoring female PTB and LPTB activity in a field setting.
Profiling
Twenty-four hours after wounding, significant shifts in the emission of several monoterpenes and monoterpenoids were observed in peach bark. These include alpha-pinene, alloocimene, beta-ocimene, beta-myrcene, camphene, linalool, and camphor. Additionally, the emission of two benzenoids, p-cymene and benzyl cyanide, increased after wounding, while benzaldehyde emissions decreased. Four of these compounds, p-cymene, linalool, benzaldehyde, and camphene, experienced similar changes in cherry. Further analysis showed that wounded peach and cherry bark only differed in the emission of p-cymene, suggesting that the emission rates of linalool, camphene, and benzaldehyde convey true information about bark damage in both peach and cherry. Further examination of the principal component loadings revealed that camphene and linalool correlate strongly with the variance between samples collected before and after wounding in both species, but do not meaningfully contribute to interspecific discrimination, while benzaldehyde emission contributes little to the discriminatory ability of all analyses. Camphene and linalool can therefore be considered likely signals of the state of physical impediments to larval entry in multiple host plants. Although shifts in the emission of benzyl cyanide, camphor, beta-myrcene, and alpha-pinene were not significant in cherry, these compounds failed to meaningfully correlate with the interspecific variance and thus should not be ignored as potential semiochemicals. Analysis of the principal component loadings likewise suggests that benzaldehyde is of low informational value insofar as discrimination between wounded and unwounded potential hosts is concerned. However, the relative abundance of a compound tends to be more important to host selection than absolute abundance and the fact that benzaldehyde emission shifts from dominating the chromatogram to become more or less equivalent to 3-hexen-1-ol in peach or nonatriene in cherry, and should probably not be ignored.
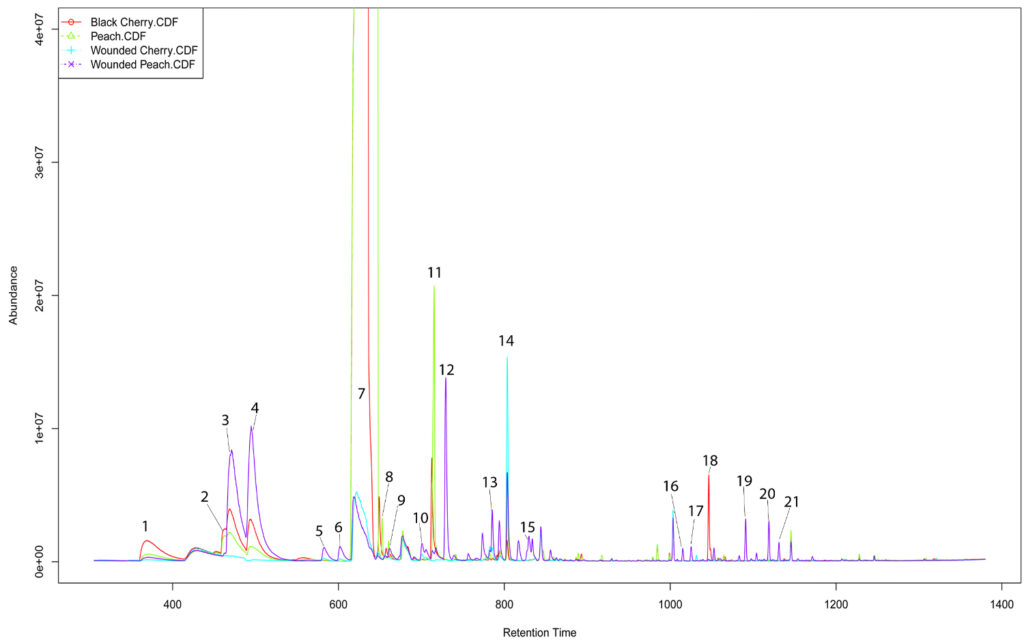
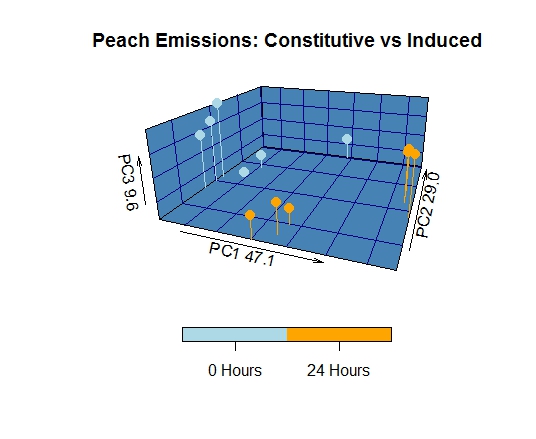
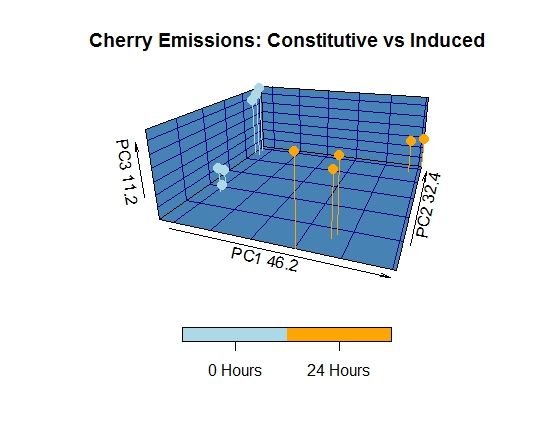
The volatile profile of cultivated peach bark/phloem, collected from a defunct orchard, differed significantly from that of wild host species. The patterns of Benzaldhyde and Benzyl Alcohol emission, as well as the abundance and diversity of GLVs and sesquiterpenes represent the most apparent variation between wild and cultivated Prunus species. GLV, benzaldehyde, and benzyl alcohol were emitted at significantly lower levels in cultivated peach trees, while sesquiterpene diversity was markedly increased. The principal component loadings provided additional support for these observations, as they showed that these compounds accounted for much of the observed variance. It is also noteworthy that mechanical damage failed to induce significant changes in the volatile profile of cultivated peach, likely as a function of the high metabolic costs associated with sesquiterpene biosynthesis.
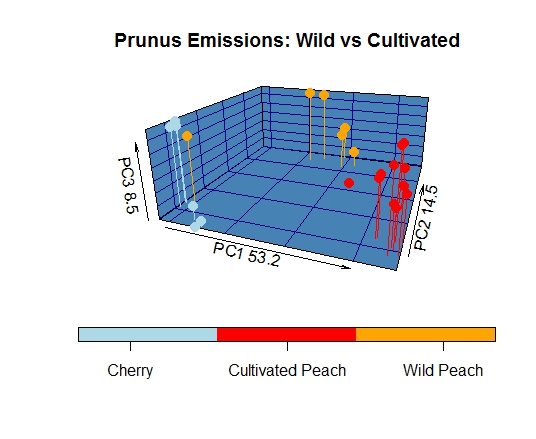
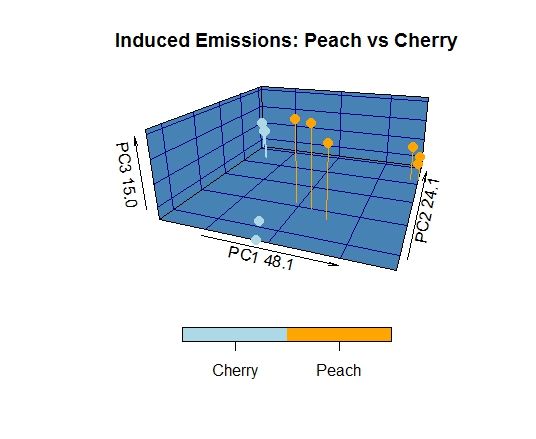
The constitutive emissions of bark/phloem collected from cultivated (non-host) apple trees differed substantially from those of Prunus species. Variability was observed in several previously identified compounds of interest, including: a-pinene, limonene, camphene, p-cymene, benzyl alcohol, and benzaldhyde. Benzyl alcohol and p-cymene were not detected in apple, while limonene was much more abundant. Camphene and alpha-pinene were increased in apple relative to host species, but relative concentrations were strongly biased towards the latter. Benzaldehyde was detected at intermediate levels, cultivated apple emitted more than cultivated peach but less than wild Prunus species. Finally, the sesquiterpene profile was almost entirely different, lacking guaiene, humulene, isocaryophyllene, and cadinene but containing longifolene and caryophyllene.
Electroantennography
It has previously been reported that a variety of plant compounds induce depolarization in the antennae of male PTBs. However, we were only able to consistently detect changes in potential associated with the presence of pheromone.
Trapping
Pheromone trapping was highly successful with each trap capturing an average of 16.3 males weekly between 21 June and 9 August 2017. However, neither the kairomone blend derived from cultivated trees, nor the blend based on wild species was able to attract female moths, and none were captured. Female activity was subsequently confirmed, indirectly, by observing the presence of early-instar larvae in the orchard.

Host Quality
We found that individuals collected from the suburban population were significantly longer than those found in the orchard (Student t-test, p = 0.02), while no discernible difference in wingspan was observed (Wilcoxon Rank-Sum Test, p = 0.11). Although major emergence events tended to occur approximately two calendar days earlier in the suburban population, this can be attributed to the slower accumulation of degree days in Faulkner County.
Larval Distribution
In areas where kairomone blends based on the emissions of orchard trees were present, new infestations were documented in forty-seven trees, 10.4% of the population. This proportion is significantly lower (Chi-Square, p = 0.0006) than the 21% and 19% of trees infested with juvenile larvae in other areas of the orchard, as shown below.
Discussion
Plants have developed a variety of chemical defenses over the course of evolutionary time. Some defensive chemicals, such as volatile phytoalexins, broadcast true information about the physiological state of a plant to the surrounding ecosystem. As a result, such semiochemicals should be utilized by those antagonists and competitors that can turn the information into reproductive advantage. Previous work determined that the PTB and LPTB (Lepidoptera: Sesiidae) lay more eggs at sites of previous infestation and mechanical wounding, respectively (Cottrell et al., 2008; Derksen et al., 2006). However, the degree to which these species utilize bark volatiles to recognize optimal oviposition sites at a distance remains unknown.
The volatile profile of cultivated peach differed substantially from that of wild trees, and showed no apparent response to mechanical damage. Green leaf volatiles were uncommon and benzaldehyde was not present in expected amounts, while an unexpected diversity and abundance of sesquiterpene compounds was observed. This observation likely stems from the fact that relatively undamaged trees were sampled from among heavily infested and infected conspecifics, as plants are known to up-regulate defensive physiology in response to the defensive emissions of neighboring plants (Maffei, 2010; Aharoni et al., 2003). Our observations align well with the explanation that defenses are primed for microbial attack, as a reduction in benzaldehyde emission is observed after wounding and sesquiterpenes have documented anti-microbial properties (Bakkali et al., 2008). The lack of an observable response to wounding is explained by the metabolic cost of constitutively emitting these sesquiterpenes, as extensive reduction is involved in their biosynthesis (Rodriguez et al., 2014).
We also note that mechanical damage stimulates the emission of several benzenoids and monoterpenoids in both black cherry and peach, suggesting that the relative emission rates of alpha-pinene, camphene, linalool, benzyl cyanide, p-cymene, and benzaldehyde convey information regarding a reduction in the physical barriers to larval entry. The utilization of these cues in the selection of oviposition sites should therefore provide a fitness benefit to the peachtree or lesser peachtree borer. However, synthetic kairomone blends proved ineffective for monitoring sesiids in the field, suggesting that bark emissions may act only as short-range ovipositional stimulants or may not convey enough information when decoupled from visual stimuli. Reliance on a single sensory input does not allow rapid validation of decisions. Many insects therefore rely on synergistic interactions between olfactory and visual cues to identify suitable host plants at relatively close range, thereby reducing predation risk or opportunity costs that may be associated with contact assessment (Goyret et al., 2007; Campbell and Borden, 2006; Raguso and Willis, 2005; Pyke et al., 1977). Hindsight leads me to the realization that we may have been too quick to focus on long-distance orientation, especially considering that we sought to examine movement within an orchard, not invasion of an orchard from the surrounding landscape.
Interestingly, the distribution of larvae in the orchard suggests that this kairomone blend may repel female PTBs. A few possibilities could give rise to this effect. Most simply, it could be a function of host quality in the area where trapping occurred, future experiments addressing this question would need to be properly controlled to rule out this possibility. This result could also arise from the qualities conveyed through the compounds emitted from our lure; we observed increased sesquiterpene concentration and diversity in the samples collected from the orchard which has been attributed to defensive priming induced by the emissions of neighboring trees. It is reasonable to posit that hosts facing microbial assault are poorer hosts. Kaur et al. (2016), for example, recently reported that fungal metabolites inhibit growth and development in the oriental leafworm moth (Spodoptera litura). Avoiding hosts with fungal infections may therefore benefit the PTB. This hypothesis is bolstered by the observation that PTBs collected from the orchard were smaller than those collected in Little Rock, suggesting that the unmanaged trees were of lower quality than suburban hosts. The repellent properties of terpenes and related compounds have also been widely documented in Lepidoptera, but many are also attractants (Binders and Robbins, 1997). Alternatively, this could result from a shift in the overall volatile profile of that orchard section. This seems somewhat unlikely, but when we consider that host finding is often mediated by highly specific emission ratios, it becomes possible. Studying the fragility of species-specific ratios could not only improve pest management strategies, but also, more fundamentally, provide insight as to the potential impacts of climate-induced shifts in volatile emissions on the foundational interactions in terrestrial ecosystems.
Educational & Outreach Activities
Participation summary:
No formal outreach activities have been performed in conjunction with this project. However, a public presentation of our findings is forthcoming.
Project Outcomes
In recent years there has been considerable interest in improving not only the efficacy of pest management regimes, but also developing more sustainable pest control strategies that allow humans to have our cake and ecosystems too. History shows the efficacy of synthetic pheromones for monitoring and mating disruption, and the past twenty years have demonstrated the role of plant-derived lures in the future of pest management (Knight et al., 2001; Natale et al., 2003; Hern and Dorn, 2004; Knight et al., 2005; Piñero and Dorn, 2009). However, the use of plant-derived semiochemicals in IPM is relatively new and faces myriad challenges. Lures derived from floral volatiles are only applicable to insects that feed as adults and may impact non-target pollinators, while it is profoundly difficult to make a lure derived from vegetative tissues stand out amongst host plants (Meagher, 2002). This report describes attempts to elucidate aspects of the chemical ecology of the sesiid peach pests with the expectation that such information will facilitate the development of more sustainable management strategies for these economically significant insects. This involved the characterization and comparison of the constitutive volatile emissions from bark of apple, black cherry, and peach, as well as the induced changes to the emissions of host species associated with mechanical damage. Although technical challenges and an extremely biased sex ratio prevented my examination of the behavioral relevance of these compounds and my kairomone blends failed to attract female moths, I observed decreases in larval abundance in areas where my lures were located. These observations provide a wealth of questions and few answers, but hint at another potential semiochemical-based pest management strategy: Deterrence through the manipulation of emission ratios.
The most important recommendation we can make is that future research should assess the fragility of the olfactory blends that mediate host finding in a well-established specialist pest system. The benefits of such research not only have the potential to reduce pest movement into agricultural fields, but also assess the robustness of these foundational interactions in the face of climate change. In this regard, Farré-Armengol et al. (2014) documented increases in the emission of floral terpenes with increased temperature, and it is unlikely that other tissues would remain unaffected.