Final report for GS17-172
Project Information
Flower thrips are worldwide pests and pose major threats as the sole vectors of tospoviruses, and affect numerous fruiting vegetable crops grown throughout the Southern region. Growers are faced with increasing challenges as thrips have developed resistance to many classes of insecticides due to a heavy reliance upon scheduled applications. In an effort to develop a sustainable integrated pest management program for pepper plants, studies have focused on exploiting naturally-occurring predators of thrips, such as the minute pirate bug Orius insidiosus.
Host plant responses to herbivory have been shown to aid in the attraction of natural enemies through the release of herbivore-induced plant volatiles. Furthermore, mixtures of multiple herbivore-induced volatiles released by different species of conspecific host plants have been shown, in some experiments, to be more attractive to flower thrips predators than volatiles produced by a single species of host plant. The effects of multiple herbivore-induced plant volatiles produced by similar conspecific host plants at different developmental stages, however, are not yet fully understood.
Determining the differences among multiple herbivore-induced plant volatiles released from pepper plants at varying maturity stages can provide insight into the ability of pepper plants to attract natural predators and, consequently, disperse pests more effectively than pepper plants of the same maturity stage. The understanding of these relationships is vital for promoting of the use of natural enemies as part of an integrated pest management program for vegetables aimed at reducing insecticide use and increasing benefits for growers, as well as the environment and community.
1. Determine the composition of single herbivore-induced plant produced by the different stages of pepper plant (vegetative vs. flowering) under direct herbivore attack.
2. Determine multiple herbivore-induced plant volatiles produced by the different stages of pepper plant (vegetative vs. flowering) while:
a. Under direct herbivore attack
b. Eavesdropping on neighboring infested plants
3. Determine if herbivore-induced plant volatiles released vary when under direct herbivore attack vs. those released in response to eavesdropping.
4. Determine if combinations of those multiple herbivore-induced plant volatiles without visual cues are more attractive to natural predators (O. insidiosus) than the single volatiles produced by the same stages using:
a. Olfactometer Y-tube testing
b. Ethovision video tracking software
5. Determine if combinations of those multiple herbivore-induced plant volatiles with visual cues are more attractive to natural predators (O. insidiosus) than the single volatiles produced by the same stages using field-cage experiments.
6. Determine the use of attractive volatiles for O. insidiosus in field conditions.
Research
Effects of insect diets on development of O. insidiosus juveniles and adult reproduction
Orius insidiosus is an omnivore and generalist predator, feeding on a wide variety of soft-bodied arthropods, eggs, plant tissue, and pollen, as well as exhibiting intra-guild predation towards earlier instar nymphs (Tommasini et al. 2002). Multiple experiments have revealed the differential prey preferences of O. insidiosus towards various species and developmental stages, including preferences for larval thrips instars over adults, and preferring WFT over other closely related flower thrips species (Baez et al. 2004, Paini et al. 2008). While these preferences commonly reflect phenotypical characteristics such as prey size and mobility, the closely related Orius majusculus has shown prey preferences towards WFT larvae over similarly immobile whitefly pupae (Arno et al. 2008). In multi-pest agroecosystems, including Florida bell pepper fields and greenhouses, the quality of available prey can also influence predatory preferences in O. insidiosus juveniles and adults. Examining the effects of insect diets commonly experienced by O. insidiosus in natural bell pepper agroecosystems in juvenile development and adult reproduction may help improve the understanding of prey preferences and improve population modeling as prey diversity and population densities fluctuate throughout the growing season.
The objectives of this study were (1) to compare the effects of insect diets on the developmental rate of O. insidiosus nymphs and (2) to compare the effects of insect diets on O. insidiosus reproduction. Cohorts of juvenile O. insidiosus were first reared to the adult stage on one of three insect diets to determine juvenile development rates. The successfully hatched progeny of reared adults for each diet were then compared to determine differences in hatched offspring. Diets consisted of juvenile stages of three naturally occurring insect species in bell pepper fields and greenhouses: WFT (F. occidentalis), Tobacco thrips (F. fusca), and the green peach aphid (Myzus persicae).
Insect Colonies
- occidentalis larvae were obtained from colonies maintained at the NFREC in Quincy, FL, USA. The colonies were established in 2018 from existing colonies donated by the United States Horticulture Research Laboratory in Fort Pierce, FL (USDA-ARS). Frankliniella fusca larvae were acquired from colonies maintained at the NFREC in Quincy, FL. The colonies were established in 2016 by a former graduate student. Both thrips species colonies were reared independently in multiple (as needed) 14 x 14 x 4.5 cm clear plastic containers (Rubbermaid, Atlanta GA) with a 6 x 6 cm ‘no-thrips’ mesh (Green-Tek, Inc., Janesville, WI) ventilation window in the lid. Store-bought green bean pods (Phaseolus vulgaris L.) soaked in a dilution containing ten percent of a six percent NaClO solution for 15 min and thoroughly rinsed and hand-dried were added to the containers three times per week at two pods per container. The containers were maintained at 25 ± 1°C, 70 ± 10% RH, and a 15:9 L: D photoperiod. M. persicae nymphs were acquired from colonies maintained at the NFREC in Quincy, FL. The colonies were established from adults collected from greenhouse pepper plants at the NFREC in Quincy, FL in 2018, and transferred to caged pepper plants in growth chamber environment at 23 ± 1°C, 40 ± 10% RH, and a 15:8 L: D photoperiod.
Wild adult O. insidiosus females were collected from wild Lagerstroemia indica L. (Family: Lythraceae) in non-disturbed areas throughout Gadsden County, FL and allowed to oviposit onto green bean pods for 24 h. Adults were then removed and pods containing the eggs of experimental cohorts were allowed to hatch for 5 days. Twenty-five first-instar nymphs (comprising one diet treatment cohort) were transferred to 14 x 14 x 4.5 cm clear plastic containers (Rubbermaid, Atlanta GA) with a 6 x 6 cm ‘no-thrips’ mesh (Green-Tek, Inc., Janesville, WI) ventilation window in the lid and maintained at 25 ± 1°C, 70 ± 10% humidity, and a 15:9 L: D photoperiod.
Experimental Protocols
Juvenile development
Cohorts received treatment diets of (1) two fresh green bean pods and F. occidentalis first and second instar larvae provided at 15/nymph/day (2) two fresh green bean pods and F. fusca first and second instar larvae provided at 15/nymph/day, (3) two fresh green bean pods and M. persicae first, second, and third instar aphid nymphs provided at 15/nymph/day or (4) only two fresh green bean pods (control diet). Green bean pods and insect diets were replenished three times per week at 2-3 d intervals, and cohort developmental stage was recorded as (1) date of at least fifty percent of cohort nymphs in the second or third instar developmental stage (early instar), (2) date of at least fifty percent of cohort nymphs in the fourth or fifth instar stage (late instar), and (3) date of at least fifty percent of nymphs at the adult stage (adult).
Adult reproduction
When cohorts were recorded as reaching the adult stage, two wild male O. insidiosus adults were introduced into the population and mated for 48-72 h. Ten female adults were then transferred by aspirator to fresh containers and treatment diets were continued. As green bean pods from the previous replenishment were rotated, old pods were placed into fresh containers and hatched nymphs were recorded every replenishment period and removed for a period of 1 w before disposing of beans. Offspring recording was terminated after two successive recording periods with zero hatched offspring observed following initial offspring eclosion. Six replications of all treatment diets were performed. Due to insufficient dietary insect colonies, F. fusca diets were recorded in the spring and summer of 2018, while F. occidentalis and M. persicae diets were recorded in the spring and summer of 2019.
Data Analysis
Effects of insect diets on juvenile development
To account for the two and three day intervals between diet replenishment and recording, time measurements were calculated as the number of days per week (7) divided by the number of recording periods per week (3) and converted into hours, which resulted in 56 h time intervals. Development time between treatments was analyzed between each developmental stage using analysis of variance
Effects of insect diets on adult reproduction
Mean total offspring hatch data between treatment diets were subjected to analysis of variance and further analyzed against each other through T-testing to determine if significant differences existed between treatment populations in reproduction. Mean offspring per recording period for F. occidentalis and F. fusca diet populations were subjected to T-testing to determine time intervals in which offspring hatch was significantly different between the two diets.
Effects of herbivore-induced plant volatiles on O. insidiosus attraction
In response to herbivore damage, plants undergo changes in their expression of defensive traits, including the release of specific volatile organic compounds (VOCs) referred to as herbivore-induced plant volatiles (HIPVs) (Karban and Baldwin 1997). Nearby natural enemies can detect these chemical signals, which provide important information to determine herbivore presence, density, and herbivore species (Dicke 1999), increasing natural enemy recruitment and activity on the infested host plant (de Morales et al. 1998). In Florida bell pepper, HIPVs aid important natural enemies, such as O. insidiosus, in locating infested host plants induced by feeding, oviposition, or eavesdropping on nearby attacked conspecifics (Arimura et al. 2002). Induced resistance theory postulates that plants incur fitness costs associated with herbivore attack and subsequent HIPV release (Cipollini et al. 2003), often varying in effects between plant species (Freundlich and Frost 2018). Additionally, the qualitative and quantitative HIPV chemical profiles released can vary significantly depending upon host plant species, herbivore species, and herbivore densities (Cai et al. 2014).
In field and greenhouse agroecosystems, multiple plant and pest species can share a common environment, and the various HIPV signals released can become mixed. While there is still much to be understood about how those mixed signals affect a common intended receiver, multiple studies have shown the attractiveness of HIPV blends over individual components (McCormick et al. 2012, Maeda et al. 2015), as well as greater attraction of Orius spp. to blends produced by combinations of multiple plant species (mHIPVs) over single plant species (sHIPVs). Furthermore, cultivars within a single plant species under attack from different herbivores can produce volatile profiles that vary in their honest signaling of herbivore density to parasitoids (Shiojiri et al. 2010). Thus, various cultivars of bell pepper may also respond differently to herbivory from pests such as the WFT, potentially altering the attractiveness of natural enemies such as O. insidiosus to mixed blends, and extending the possibility of manipulation strategies if blends of multi-cultivar HIPVs are more attractive to natural enemies than single-cultivar blends.
The following set of experiments investigated the attractiveness of various HIPV blends produced by WFT-infested bell pepper cultivars to O. insidiosus. For the first experiment, an indoor cage experiment tracked O. insidiosus populations exposed to various induced bell pepper combinations over time. For the second set of experiments O. insidiosus olfactory responses to headspace volatiles from cultivar combination were analyzed in a series of dual-choice Y-tube bioassays. The objective of these experiments was to determine if differences in attractiveness exist between HIPVs of similar and multi-cultivar pepper combinations to O. insidiosus adults.
Insects and Plants
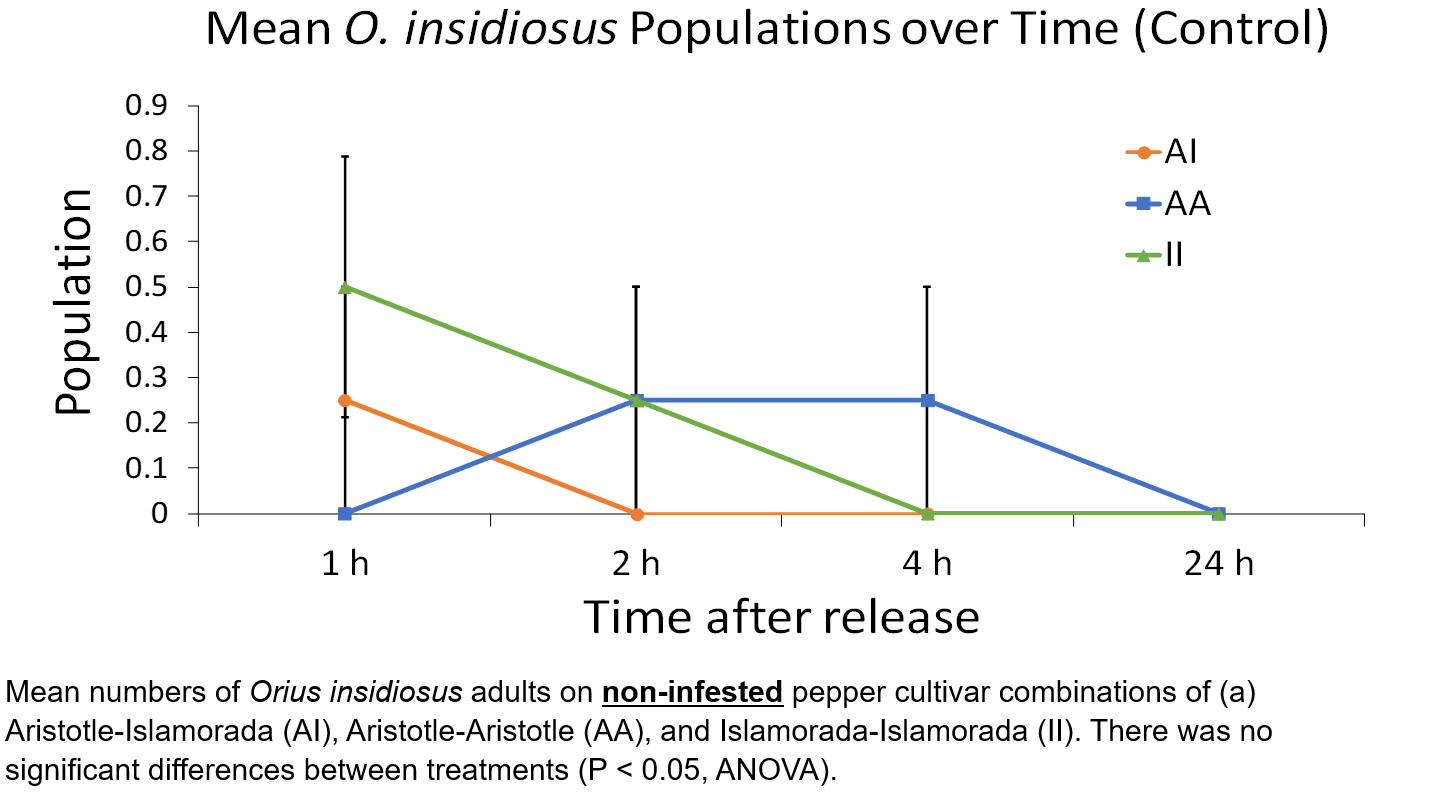
Cultivar combination attraction to O. insidiosus populations over time in a cage environment
- occidentalis larvae were acquired from existing colonies at the NFREC in Quincy, FL. The colony was established in 2018 using adults donated from existing colonies maintained at the United States Horticultural Research Laboratory in Fort Pierce, FL. Colonies were maintained in 14 x 14 x 4.5 cm clear plastic containers (Rubbermaid, Atlanta GA) with a 6 x 6 cm ‘no-thrips’ mesh (Green-Tek, Inc., Janesville, WI) ventilation window in the lid. Store-bought green beans (Phaseolus vulgaris L.) soaked in a solution containing 10 percent of a 6.0 percent NaClO and thoroughly rinsed and dried were added to the containers three times per week. The containers were maintained at 25 ± 1°C, 60 ± 10% RH, and a 15:9 L: D photoperiod.
- insidiosus adults were collected as fourth and fifth instar nymphs from wild, undisturbed crimson clover flowers (Trifolium incarnatum L.) in Gadsden County, FL in April, 2019 and reared to the adult stage in inflorescence of the collected host plant containing naturally occurring flower thrips populations and one hundred artificially introduced F. occidentalis first and second instar larvae per day to prevent mixing of previously-exposed and naïve O. insidiosus populations. Colonies were maintained in 42 x 28 x 14 cm clear plastic containers (Rubbermaid, Atlanta GA) with red lids cut into 27 x 14 cm ventilation windows covered with ‘no-thrips’ mesh at 21 ± 1°C, 50 ± 10% RH, and a 15:9 L: D photoperiod. O. insidiosus populations were 3-10 d at the adult stage during experiments.
Aristotle 3XR hybrid and Islamorada sweet bell peppers cultivars were grown in a greenhouse environment at the NFREC in Quincy, FL. All plants were in the vegetative stage of development with between four and six true leaves and 5-8 w. Due to occurrences of plant infestation of small thrips and aphid populations, all pepper plants were exposed to five F. occidentalis larvae prior to experimentation after reaching the experimental development between 48-72 h prior to use to reduce the possibility of naïve and previously-induced plants becoming mixed in experiments, which may affect host plant volatile synthesis and release time, as well as qualitative and quantitative chemical profiles (Pare and Tumlinson 1999).
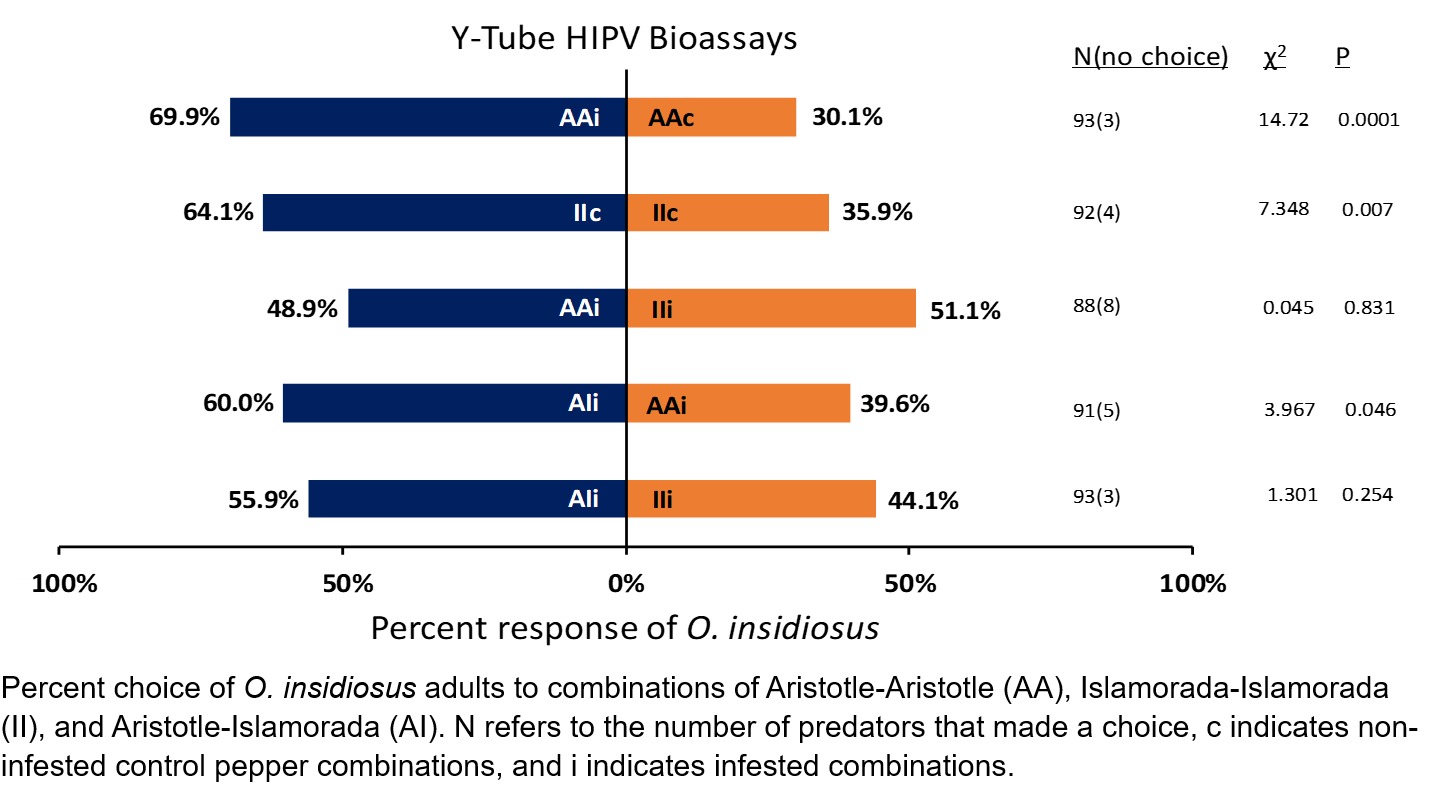
Olfactory responses of O. insidiosus in Y-tube olfactomer bioassays
- occidentalis larvae were acquired from existing colonies at the NFREC in Quincy, FL. The colony was established in 2018 using adults donated from existing colonies maintained at the United States Horticultural Research Laboratory in Fort Pierce, FL. Colonies were maintained in 14 x 14 x 4.5 cm clear plastic containers (Rubbermaid, Atlanta GA) with a 6 x 6 cm ‘no-thrips’ mesh (Green-Tek, Inc., Janesville, WI) ventilation window in the lid. Store-bought green beans (Phaseolus vulgaris L.) soaked in a solution containing 10 percent of a 6.0 percent NaClO and thoroughly rinsed and dried were added to the containers three times per week. The containers were maintained at 25 ± 1°C, 60 ± 10% RH, and a 15:9 L: D photoperiod.
- insidiosus adults were collected as fourth and fifth instar nymphs from undisturbed crape myrtle flowers (Lagerstroemia L.) in Gadsden County, FL in summer, 2019 and reared to the adult stage in inflorescence of the collected host plant containing naturally occurring flower thrips populations and one hundred artificially introduced F. occidentalis first and second instar larvae per day to prevent mixing of previously-exposed and naïve O. insidiosus populations. Colonies were maintained in 42 x 28 x 14 cm clear plastic containers (Rubbermaid, Atlanta GA) with red lids cut into 27 x 14 cm ventilation windows covered with ‘no-thrips’ mesh at 21 ± 1°C, 50 ± 10% RH, and a 15:9 L: D photoperiod. O. insidiosus populations were 3-10 d at the adult stage during experiments.
Aristotle 3XR Hybrid and Islamorada sweet bell peppers cultivars were grown in (pot size) in a greenhouse environment at the NFREC in Quincy, FL. All plants were in the vegetative stage of development with between four and six true leaves. Due to occurrences of minimal greenhouse infestation of thrips and aphid species, all pepper plants were exposed to five F. occidentalis larvae prior to experimentation after reaching the experimental development stage 48-72 h prior to use to minimize the possibility of naïve and previously-induced plants becoming mixed, which may affect host plant volatile synthesis and release time, as well as qualitative and quantitative chemical profiles (Pare and Tumlinson 1999).
Treatments to test attractiveness to O. insidiosus
Cultivar combination attraction to O. insidiosus populations over time in a cage environment
Three F. occidentalis-infested combinations of (1) Aristotle-Aristotle (AAi), (2) Islamorada-Islamorada (IIi), and (3) Aristotle-Islamorada (AIi) and three non-infested control combinations of (1) Aristotle-Aristotle (AAc), (2) Islamorada-Islamorada (IIc), and (3) Aristotle-Islamorada (AIc) were used in cage experiments. Infested treatment combinations were exposed to 30 F. occidentalis first and second instar larvae (15 larvae per plant) 3 h prior to O. insidiosus exposure and placed into the experimental cage environment. An approximately 1:1 ratio of 25 O. insidiosus male and female adults were transferred together into a 14 x 14 x 4.5 cm clear plastic container (Rubbermaid, Atlanta GA) with a lightly misted 7.5 x 7.5 cm paper tower square containing distilled water and a 6 x 6 cm ‘no-thrips’ mesh (Green-Tek, Inc., Janesville, WI) ventilation window in the lid and starved for 3 h during the pepper infestation period.
Olfactory responses of O. insidiosus in Y-tube olfactomer bioassays
Three F. occidentalis-infested combinations of (1) Aristotle-Aristotle (AAi), (2) Islamorada-Islamorada (IIi), and (3) Aristotle-Islamorada (AIi) and two non-infested control combinations of (1) Aristotle-Aristotle (AAc) and (2) Islamorada-Islamorada (IIc) were used olfactometer Y-tube assays. Infested pepper combinations were exposed to 30 F. occidentalis first and second instar larvae (15 larvae per plant) 3 h before O. insidiosus exposure. Twenty-four O. insidiosus female adults were placed into a plastic quart-sized Ziploc bag with a lightly misted 7.5 x 7.5 cm paper towel square containing distilled water and starved for 3 h during the pepper infestation period.
Experimental Environment
Cultivar combination attraction to O. insidiosus populations over time in a cage environment
Cages were constructed from white PVC pipes connected to form a 104 cm3 frame and wrapped in sewn mesh to prevent insect escape. A velcro lining around the borders of two opposite ends of the cage allowed experimenter access to the arena. The mesh cage sat on a metal shelving unit and raised 7 cm off the ground and was surrounded by white foam core to prevent visual bias from the surrounding environment. A 60W daylight bulb was fixed at the top center of the mesh ceiling. Cages contained three equidistant treatment pepper combinations located 30 cm from each treatment combination in an approximately circular fashion.
Olfactory responses of O. insidiosus in Y-tube olfactomer bioassays
Assays were conducted in an experimental environment constructed using a Y-tube (12 cm common tube, 10 cm arms, 2.5 cm internal diameter; Sigma Scientific, Gainesville FL) suspended vertically by a steel clamp attached to a support stand and surrounded by a white foam core on three sides with a 60W equivalent daylight LED bulb overhead. Pepper plant treatment combinations were placed in one of two plastic bags for headspace volatile collection and secured with plastic zip ties.
Experimental Protocols
Cultivar combination attraction to O. insidiosus populations over time in a cage environment
Cage experiments were conducted at room temperature. One plastic container containing starved O. insidiosus adult males and females was placed into the center of the arena and the lid removed. Adults were exposed to cages treated with (1) F. occidentalis-infested combinations of Aristotle-Aristotle, Islamorada-Islamorada, and Aristotle-Islamorada. Initial population release was performed at 1200. Numbers of O. insidiosus adults present on plants of each treatment combination were recorded following initial release after (1) 1 h, (2) 2 h, (3) 4 h, and (4) 24 h. Counting was done by opening access windows and inspecting each plant while remaining in the arena. Access windows were securely sealed after each inspection to prevent escape. One 24 h cage trial was counted as one replicate, and four replicates were performed. Pepper combinations were rotated each replicate to prevent directional bias.
Olfactory responses of O. insidiosus in Y-tube olfactometer bioassays
Y-tube bioassays were conducted at room temperature. Twenty-four O. insidiosus females were individually assayed per replication and allotted 5 min to choose one of the two arms containing headspace volatiles from two treatment pepper combinations by passing at least halfway through the arm. If no choice was made, the trial was recorded as ‘no choice.’ In five separate assays, twenty-four individual O. insidiosus females were presented with headspace collections from two-plant combinations of (1) infested and non-infested Aristotle plants (AAi vs. AAc) as a positive control, (2) infested and non-infested Islamorada plants (IIi vs. IIc) as a positive control, (3) infested Aristotle and infested Islamorada plants. (AAi vs. IIi), (4) infested Aristotle and infested Aristotle-Islamorada mixed plants (AAi vs. AIi), and (5) infested Islamorada and infested Aristotle-Islamorada mixed plants (IIi vs. AIi). Four replications of all dual-choice experiments were conducted (n = 96). Every six assays the Y-tube and odor source was rotated to opposite ends to control for directional bias, and airflow source was alternated each replication. Assays were conducted between the hours of 1200-1600. After one replication, glassware was soaked in hot water containing Sparkleen soap, rinsed with hot tap water followed by acetone, and allowed to dry for 2-3 d in an oven at 50 degrees Celcius.
Data Analysis
Cultivar combination attraction to O. insidiosus populations over time in a cage environment
Mean O. insidiosus populations for each pepper combination were calculated over each of the time intervals. Populations on treatment combinations during the same interval were analyzed using a one-way ANOVA to determine if there were significant differences in attraction between the three treatment combinations. Mean population differences on treatment combinations between time intervals were subjected to a one-way ANOVA to determine if there were significant rates of change in treatment densities over time.
Olfactory responses of O. insidiosus in Y-tube olfactometer bioassays
Response data minus ‘no choice’ data for all replications were pooled, and a chi square analysis performed with an expected probability of 0.5 for each treatment to determine if differences in attraction between presented choices existed. ‘No choice’ data was calculated as a percentage of sample size for each treatment.
Effects of insect diets on O. insidiosus development and reproduction
Results
Effects of insect diets on juvenile development
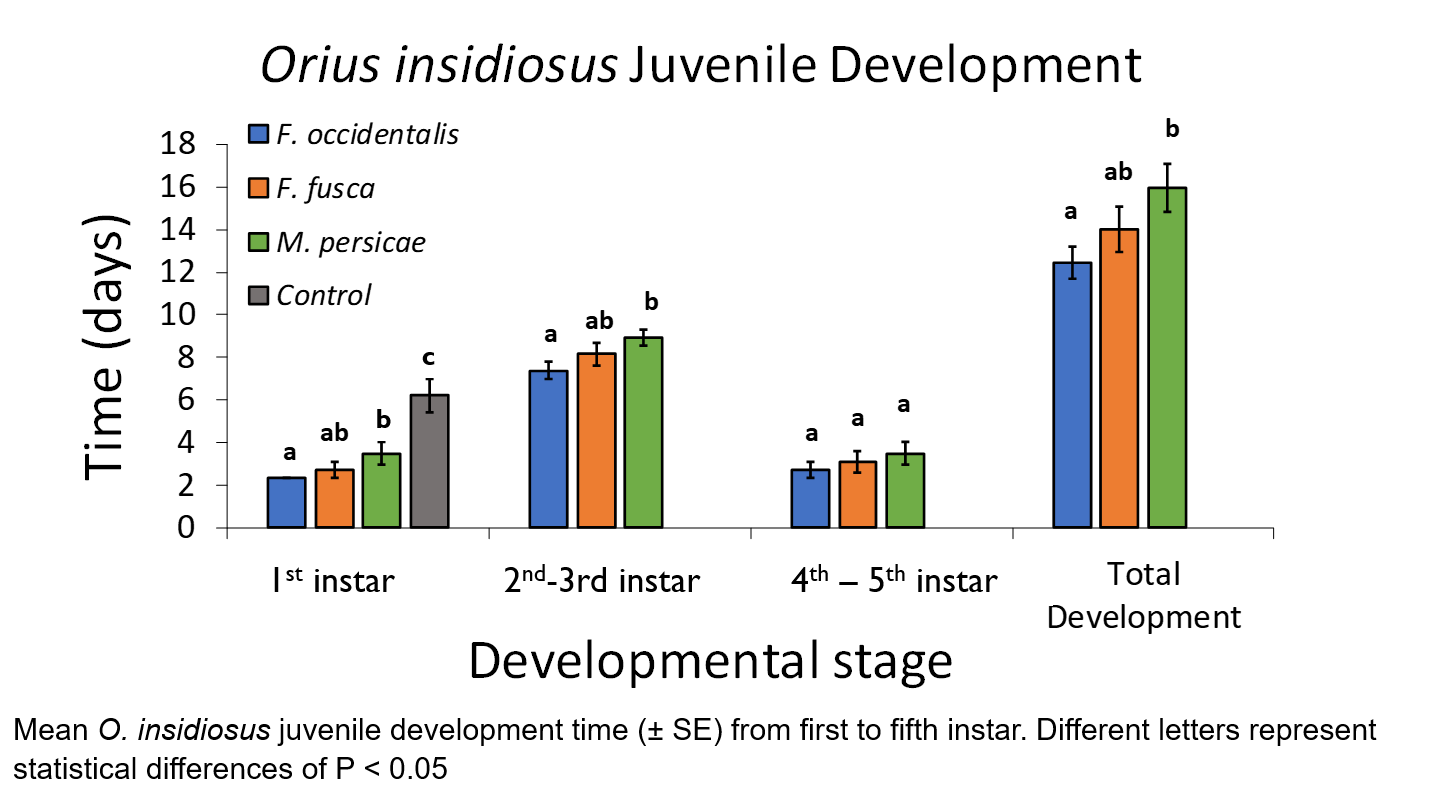
Juveniles reared on control diets of green-beans-only did not develop beyond the early nymphal stage, and thus were not used in statistical analysis. All treatment insect diets resulted in juvenile development to the adult stage. No treatment population spent a significantly different amount of time within any developmental stages (table). However, F. occidentalis treatment juveniles developed to the late instar stage significantly earlier than M. persicae populations in overall development time (t = 2.91, df = 5, P = 0.03), but this difference was not significant between F. occidentalis and F. fusca diets (t = 1.17, df = 5, P = 0.30). There were no significant differences in rate of change to successive stages, and there were no significant differences in total time to adult development.
Effects of insect diets on adult reproduction
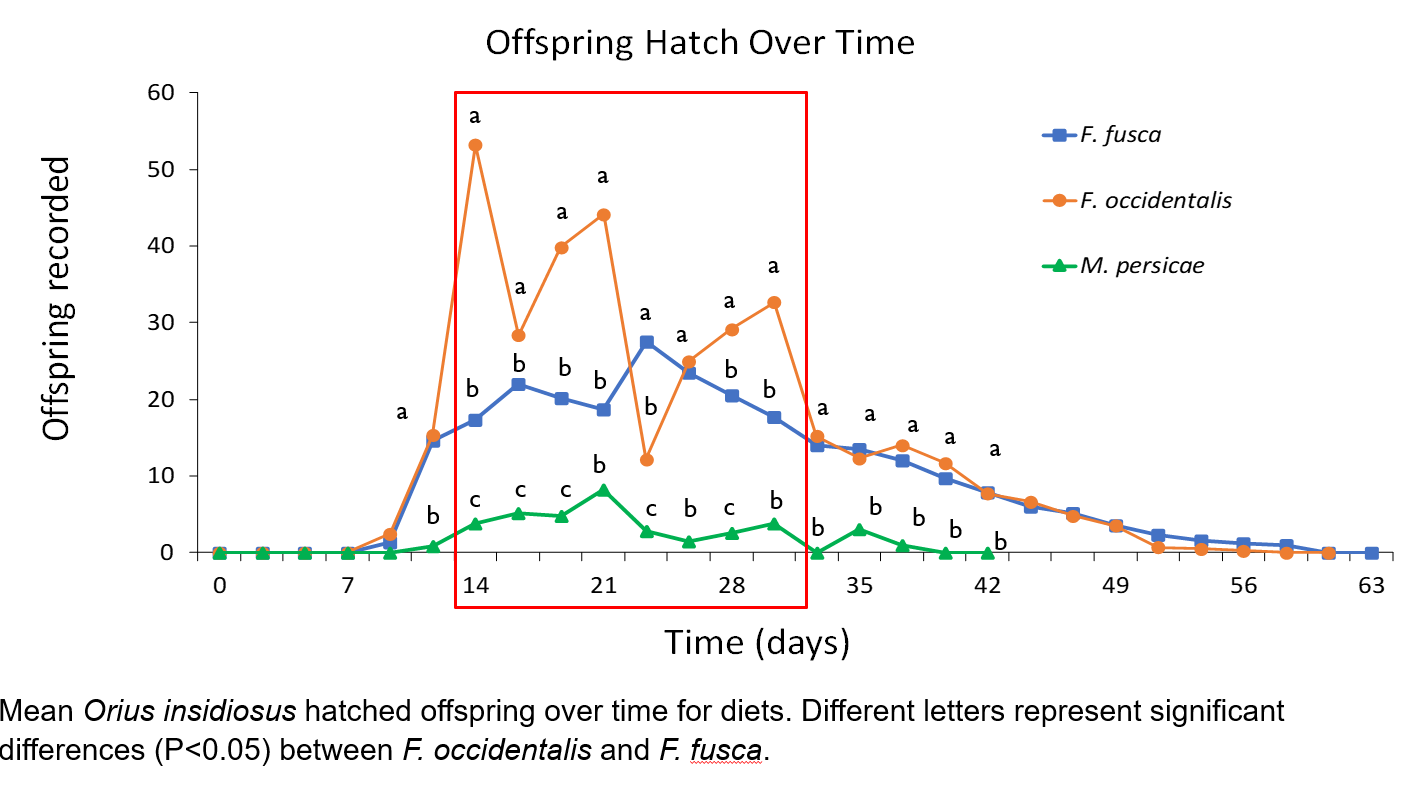
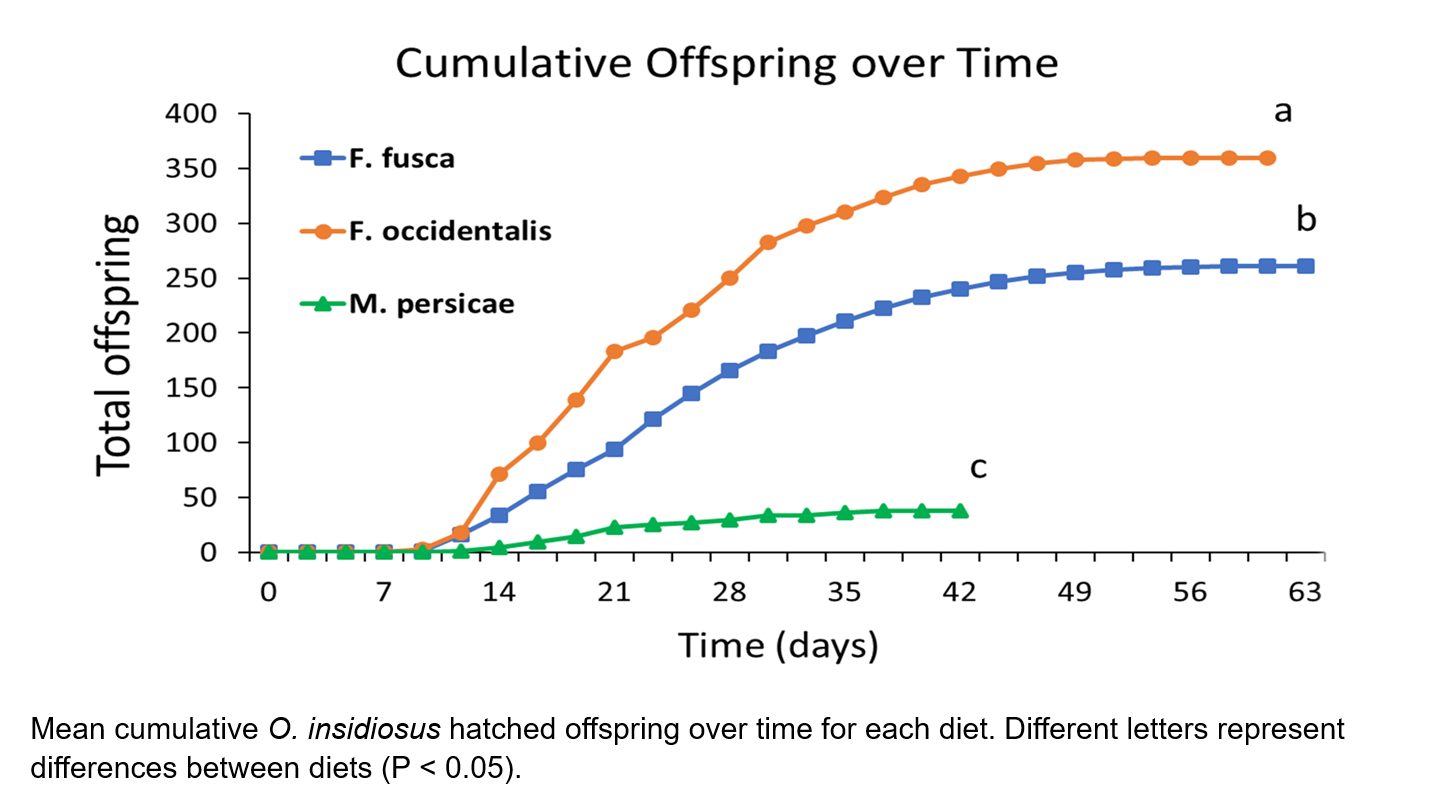
Mean offspring hatched during individual recording periods from eggs produced by F. occidentalis treatments were significantly greater than F. fusca diets for recording periods 7-10 and 13-14, or between days 14-21 and 28-30.3, respectively. During recording period 11 (day 23.3), mean F. fusca treatment offspring were significantly greater than F. occidentalis treatments. ANOVA between mean total hatched offspring showed differences between treatments, and further analysis revealed significantly greater total F. occidentalis treatment offspring compared to F. fusca diet offspring (t = 4.12, df = 5, P = 0.009) as well as greater total F. fusca diet offspring compared to M. persicae diet offspring (t = 7.30, df = 5, P < 0.001).
Discussion
The results from these experiments agree with previous research showing WFT to be a preferred prey species over other flower thrips or non-related prey (Baez et al. 2004). While total development time to the adult stage was not affected by treatment diet, earlier nymphal development to the late instar stages from a diet of WFT is important in prey handling and overcoming defensive behaviors such as abdominal jerking (Bakker and Sabelis 1987).
The greater number of total hatched offspring from O. insidiosus females feeding on WFT larvae may reveal differences in nutritional quality necessary for adult reproduction, but not necessarily juvenile growth and development. Indeed, diets of F. occidentalis and M. persicae juveniles resulted in similar adult developmental times, but total hatched offspring were significantly greater and occurred over a longer period with adults fed F. occidentalis diets. Previous research has revealed the relatively low energy provided by aphid diets for arachnids (Toft 1995), which may explain differences in offspring populations, as energy requirements are increased for gravid females. However, we observed greater dispersal behaviors of M. persicae populations from green bean pods during diet replenishment, as well as the presence of honeydew excretions on pods, both of which may reduce predator-prey interaction and successful oviposition and hatching in areas of green bean pods covered in honeydew. Thus, nutritional quality in development and offspring production may be better compared between WFT and tobacco thrips larval diets, whose adults and larval stages are commonly located within the flowers of their host plant (Hansen et al. 2003, Kahn et al. 2005). The significantly higher total offspring of both Frankliniella spp. compared to M. persicae offspring populations, as well as significantly greater total offspring from F. occidentalis diets compared to F. fusca diets, implicate a hierarchy of preferable diets that agree with the foraging behaviors of O. insidiosus, including primary concentration within flowers commonly shared by Frankliniella spp. as well as prey preferences towards WFT species. (Hansen et al. 2003, Baez et al. 2004)
While individual, consistent insect diets may significantly affect total offspring of predatory populations, more natural environments, including fields and greenhouses, commonly experience multi-pest agroecosystems with regular fluctuations in prey densities and diversity (Albajes and Alomar 1999, Bouagga et al. 2018), all of which may vary in their effects on development and reproductive success when consumed as part of a polyphagous diet. Therefore, accurate assessment of prey density and diversity through scouting is critical to better understanding how multi-pest populations may affect predator and prey population models over time to determine if ratios will meet the criterion for suppression and control (Funderburk 2000).
Effects of HIPVs on O. insidiosus attraction
Results
Cultivar combination attraction to O. insidiosus populations over time in a cage environment
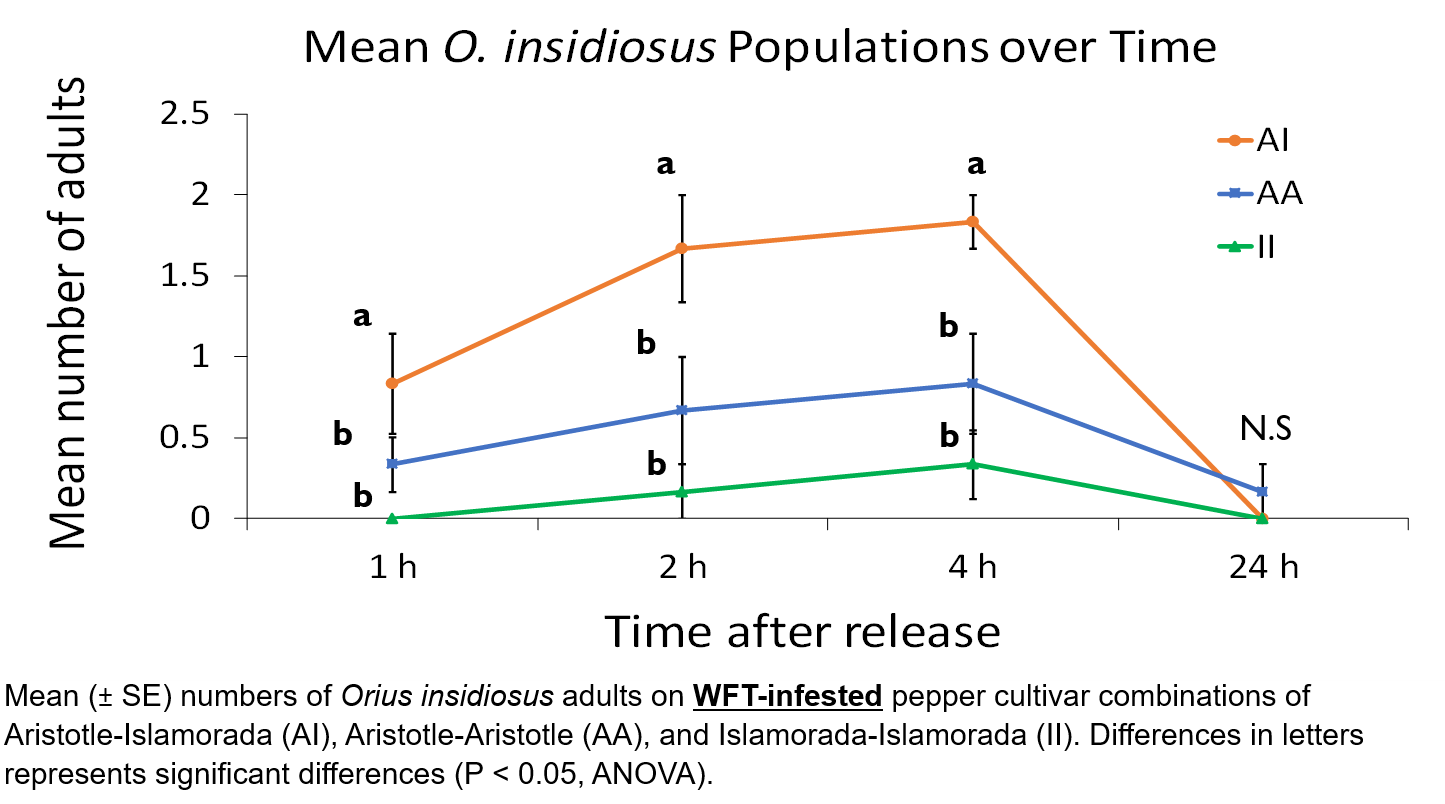
There were significant differences in O. insidiosus populations between treatment plants at 1 h (F = 4.77, df = 2, P = 0.024), 2 h (F = 7.00, df = 2, P = 0.007), and 4 h (F = 10.5, df = 2, P = 0.001). Further analysis revealed that population densities were significantly higher on Aristotle-Islamorada plant combinations than Aristotle-Aristotle plant combinations at 1 h (t = 3.16, df = 5, P = 0.025), 2 h (t = 3.87, df = 5, P = 0.011), and 4 h (t = 2.74, df = 5, P = 0.04). Population densities were also significantly higher on Aristotle-Islamorada plant combinations than Islamorada-Islamorada plant combinations at 1 h (t = 2.71, df = 5, P = 0.042), 2 h (t = 4.39, df = 5, P = 0.007), and 4 h (t = 4.39, df = 5, P = 0.007). There were no significant differences in O. insidiosus population densities between treatments at 24 h (F = 1.0, df = 2, P = 0.39), with all treatment densities reduced to at or near zero. Additionally, there were no significant differences between Aristotle-Aristotle and Islamorada-Islamorada populations.
Olfactory responses of O. insidiosus in Y-tube olfactometer bioassays
In the first two HIPV olfactometer assay experiments, O. insidiosus adults preferred Aristotle-Aristotle infested (χ2 = 14.72, df = 1, P = 0.0001) and Islamorada-Islamorada infested plants (χ2 = 7.35, df = 1, P = 0.0068) over Aristotle-Aristotle and Islamorada-Islamorada non-infested plants, respectively. In the third assay, adults showed no significant preference towards Aristotle-Aristotle infested or Islamorada-Islamorada infested plants (χ2 = 0.0455, df = 1, P = 0.83). In the fourth and fifth assays, adults showed a significant preference towards infested Aristotle-Islamorada plants over infested Aristotle-Aristotle plants (χ2 = 3.97, df = 1, P = 0.046), but showed no preference between Aristotle-Islamorada infested and Islamorada-Islamorada plants (χ2 = 1.30, df =1, P = 0.254).
Discussion
Florida bell pepper growers face increasing challenges from economically important pests such as the WFT, and the improvement of biological control strategies through understanding the effects of HIPVs in natural enemy attraction, in conjunction with other effective and vertically integrated IPM techniques dedicated to WFT control, offers safe and sustainable alternatives over regular broad spectrum insecticide applications (Weiss et al. 2009). The experiments conducted reveal differences in attractiveness that occur not only between plant species, but between cultivars of conspecifics. Furthermore, the blends produced from multi- cultivar combinations result in consistently higher O. insidiosus population densities compared to individual combinations during the first four hours of exposure in cage trials. The results reveal the possibility of predatory behavioral manipulation in natural environments to exploit O. insidiosus search behaviors and increase predatory populations and prey-interactions in WFT-infested fields. The resulting increases in juvenile and adult populations would contribute to more effective O. insidiosus to WFT ratios necessary for biological control of the pest (Funderburk et al. 2009).
While the initial results and implications of these experiments justify further research, the effects of HIPV blends on predator attraction in field trials present unique challenges. Qualitative and quantitative HIPV profiles can vary significantly within plants and are influenced by a variety of biotic and abiotic conditions, including herbivore species, herbivore density, attack from multiple species, soil humidity, fertilization, and temperature (Scala et al. 2013). Additionally, thousands of C. annuum cultivars are in use around the world (Prohens and Nuez 2008), though in the state of Florida it is estimated that 4-5 cultivars account for 80-90 percent of state-wide commercial production (McAvoy and Ozores-Hampton 2017). Consequently, the exploitation of O. insidiosus search behaviors through the manipulation of multi-cultivar HIPV blends may require preliminary attraction experiments, volatile profile analysis, and situational consideration of factors such as climate, cultivars, and pest diversity within the experimental environment.
Improved prey-location through multi-level behavioral manipulation from both long-range host plant HIPV signaling followed by close-range contact cues may improve predatory effectiveness in agroecosystems at risk of thrips vector populations when combined with proper monitoring techniques and conservational approaches to preserve natural O. insidiosus and competing thrips populations. However, there is still much research necessary to determine the differences in chemical profiles and interactions of pepper cultivar HIPVs under the relatively wide range of biotic and abiotic conditions experienced in natural environments. Additionally, honest signaling is an important component of behavioral manipulation, and strategies utilizing HIPVs and prey contact cues must continue to evaluate the effects of induced chemical profiles and their attractiveness under varying circumstances such as herbivore species to determine if increased attractiveness can be maintained throughout the growing season.
Educational & Outreach Activities
Participation summary:
From this project we plan to have 3 journal articles submitted to peer-reviewd journal by 2020. One has been submitted this summer to Entomologia Experimentalis and Applicata. Edward Traczyk presented his research at four different scientific meetings. Edward participate to the 4-H outreach activity of the lab (estimated 250 students), to field days dedicated to vegetable pest management, and to the 4-H insect club of Leon County.
Presentations Title (extension talks during field days):
4/11/2018: The state of IPM technologies in vegetable and row crops. Certified Crop Advisor training. 64 attendees. Online training.
01/16/2018: The state of IPM technologies, 2018 Agronomic In-Service Training, Quincy FL. (31 attendees
12/7/2017 Management of insect pests in Tomatoes. Gadsden county tomato Forum. 80 attendees
10/13/2017 Management of Whitefly in north Florida. Tomato field day, Quincy FL. 60 attendees.
1/17/2017 – The state of IPM technologies, 2017 Agronomic In-Service Training, Quincy FL. (31 attendees)
Presentation at scientific conferences:
Traczyk E., Martini X. Poster Foraging behavior of a minute pirate bug under starvation and feeding. Annual meeting of the Southeastern branch of the Entomological society of America. March 4-7 2018, Orlando FL. First place of the Master student poster competition.
Traczyk E., Martini X. Symposium. Foraging behavior of a minute pirate bug under starvation and feeding. Florida Entomological Society Meeting. July 22-25, 2018, St. Augustine, FL.
Traczyk, E., Poster Funderburk J., McAuslane, H., and Martini, X. Predatory search behaviors of a minute pirate bug, Orius insidiosus (Hemiptera: Anthocoridae), in response to thrips contact cues. International Society of Chemical Ecology 2019 Annual Meeting. Atlanta, GA June 2- 6, 2019.
Traczyk E., Martini X., Funderburk J., McAuslane H. Poster Effects of western flower thrips contact cues on the foraging behaviors of a minute pirate bug, Orius insidiosus. Annual Meeting of the Southeastern Branch of the Entomological society of America. March 3-6, 2019, Mobile, AL. Second place of the Master student poster competition
Project Outcomes
These effects are not believed to be limited to O. insidiosus for any reason, and may serve as useful knowledge to a broader range of natural enemies. Combined with other IPM strategies such as push-pull systems currently in place, contact cues and HIPV manipulation may provide a powerful tool in combating western flower thrips and improving biological control early in the growing season. HIPV manipulation from two alternating same-species row crops appears to be a relatively cost-efficient solution, and these findings warrant future research to determine if growing strategies using the methods analyzed result in similar successes in natural environments. If field results reflect our findings, the increased attractiveness of O. insidiosus to these volatile blends may help in maintaining O. insidiosus to thrips population ratios below the established economic injury levels, resulting in a lower demand for insecticide application and their associated costs, which would also lower risk of induced resistance and natural enemy and competitor species elimination from insecticide use.
As a MS student, this research not only helped develop a broad understanding of integrated pest management in agroecosystems, but the more focused and emergent fields of behavioral manipulation through exploiting the multi-trophic chemical ecology between plants, pests, and predators. We believe this field of study is critical in future IPM strategy development, and results show the potential of volatile organic compound blends in behavior manipulation of important biological control agents. We believe these strategies are suitable candidates for sustainable approaches to agricultural pest management for the western flower thrips and their natural enemies, and may result in lowered insecticide usage and costs, both of which reflect the overall goals of integrated pest management and sustainable agriculture.
Future research involving other cultivars and field trials will be necessary to assess the effectiveness of volatile blends in attraction of predatory bugs in a natural environment. Additionally, IPM strategies such as push-pull systems may want to be incorporated into these strategies, and will require further testing to determine how those strategies integrate. In areas where multiple minute pirate bug species co-occur, it will need to be determined how these other species will respond to similar volatile blends and how those behaviors may effect results. We plan to capture and analyze these volatile blends in the future, as well as determine if those blends introduced separately have an additive or potentially multiplicative effect, as these experiments only introduced the mixed blends at one time.