Final report for GS23-281
Project Information
Cassava is a root crop grown throughout the tropics, often in intercropping systems with legumes. Many Americans in the Southern Region consume cassava regularly, especially those of Caribbean, Latin American, and African descent. However, while subtropical regions such as Florida and Puerto Rico have a suitable climate and environment for cassava production, most cassava eaten in the US is imported. To improve local food security and access to culturally valuable foods, this project aims to investigate the potential of intercropping cassava with legumes to form a resilient cropping system for local farmers with potential benefit in larger-scale systems. Four grain legumes (peanut, soybean, cowpea, and Bambara groundnut) were tested for compatibility with cassava in North-Central Florida. Factors included effects of intercropped legumes on weed pressure and nutrient cycling in cassava-legume systems.
Objectives of the research were as follows –
- Assess the capacity of four grain legumes (cowpea, peanut, soybean, and Bambara groundnut) to suppress weeds in the presence and absence of manual weeding for a cassava intercrop.
- Investigate the mechanisms of competition between cassava and legumes using a container experiment with artificial barriers to separate above and below ground competitive effects.
- Assess cassava productivity of four cassava-legume intercropping systems as a function of legume residues as green manure after legume harvest.
Research
Objective 1
Experimental design
Experiments will be carried out at the University of Florida Plant Science Research and Education Unit (PSREU) in Citra, FL, a location at the northern end of the range where cassava may be able to be cultivated in Florida. Cassava and four grain legumes (peanut, soybean, cowpea, and Bambara groundnut) will be grown in monoculture and intercropped for a total of 9 treatments (sole cassava, sole peanut, sole soybean, sole cowpea, sole Bambara groundnut, cassava + peanut, cassava + soybean, cassava + cowpea, and cassava + Bambara groundnut) in RCBD with four replications. The experiment will be run in each of the two years of this project. All treatments will be planted in early May. Legumes will be harvested at maturity, 3-4 months after planting, and cassava will be harvested before the first frost, in late November or early December. Yield, above-ground biomass, and below-ground biomass will be measured for each crop at harvest.
Yield and biomass
Yield and biomass data will be analyzed in SAS to quantify the productivity and competitiveness of each crop in monoculture and intercropping. Sole and intercropped yields will be used to calculate land equivalent ratio (LER), a standard measure of intercropping productivity, and area-time equivalent ratio (ATER), a modification of LER that accounts for the different growth durations of cassava and legumes in these systems (Mutsaers et al., 1993). Cassava-legume intercropping systems that demonstrate high yields and positive LER and ATER values in this environment may be recommended to Florida farmers. If LER and ATER values indicate that cassava-legume intercropping is not beneficial in this environment, cassava monocultures may be more suitable.
Biomass data will be used to calculate harvest index (HI) for each crop. Part of cassava's response to competition in intercropping is to change partitioning of assimilates to below-ground biomass (yield) and above-ground biomass (canopy), so quantifying the impact of each cropping system on cassava HI will help identify which legumes, if any, are suitable for intercropping with cassava in the US.
Qualitative assessment
Since cassava has not been widely studied in the US, information on sprouting times, pest and disease pressure, and growth habit will be useful for developing best practices for cassava in this environment. To this end, observations of the performance of cassava in Citra, FL will be recorded for use in extension documents.
Objective 2
Experimental design
To assess the effect of legumes on weed management and nutrient cycling when intercropped with cassava, the experimental design described for Objective 1 will be augmented to create two factorial experiments at PSREU in Citra, FL.
For weed management, a 9 x 2 factorial experiment will be conducted, with factor A (9 levels) being the cassava and legume cropping systems described for Objective 1, and factor B (2 levels) being the presence or absence of weeding (at 3 and 6 weeks after planting). Weed abundance and biomass will be measured in each plot using a 1 x 1 meter quadrat at legume harvest and at cassava harvest.
For nutrient cycling, a 9 x 2 factorial experiment will be conducted, with factor A (9 levels) being the cassava and legume cropping systems described for Objective 1, and factor B (2 levels) being the removal of legume residues after legume harvest or the retaining of legume residues on the soil surface. Four litterbags of legume residues will be placed on each plot where residues are retained, and will be removed after 2 weeks, 4 weeks, 8 weeks, and at cassava harvest to measure residue decomposition rates. Biomass samples from collected litterbags and from fresh legume residue will be sent to the UF Forage Evaluation Lab to measure nitrogen concentrations in the residues at each timepoint.
In summary, to accomplish both Objective 1 and Objective 2, 2 RCBD experiments with 18 treatments each will be planted at PSREU in each of two years.
Yield
As with Objective 1, yield for each crop and LER and ATER for each intercropping system will be compared to assess the productivity of each system and the effect of weed and nutrient management techniques. A mixed model approach will be used for analysis in SAS, with legume, intercropping status, weeding, and residue management as fixed effects, and block and year as random effects.
Our first hypothesis is that weeding at 3 and 6 weeks will have a negative effect on LER, meaning that cassava, which grows slowly initially, benefits from the ability of legume intercrops to overshadow and suppress weed growth in the first few months after planting. Reduced need for weeding in intercropping systems compared to monocultures would therefore be a benefit of cassava-legume intercropping. Carrying out this experiment with four different legumes will allow us to identify which legumes, if any, are most useful for weed management in intercropping.
Our second hypothesis is that retaining legume residues on the soil after legume harvest will have a positive effect on LER, meaning that cassava, which continues growing for several months after intercropped legumes are harvested, may benefit from the mineralization of nitrogen and other nutrients in legume residues, particularly in the subtropical Florida environment where sandy soils, high temperatures, and high precipitation lead to rapid decomposition of plant residues. Carrying out this experiment with four different legumes will allow us to identify which legumes, if any, have the most beneficial residues for nutrient cycling in intercropping.
Secondary measures
Measuring weed abundance and biomass allows confirmation of the effectiveness of the weeding treatment in reducing weed pressure, and comparison of weed pressure between all treatments. We expect that weed abundance and biomass, particularly weed biomass, will correlate negatively with cassava and legume yields.
Litterbags in the retained residue treatments help determine if legume residues mineralize quickly enough for their nutrients to be accessible to intercropped cassava in these systems and in this environment. If there is a benefit to residue retention but litterbags indicate that legume residues are not mineralizing before cassava harvest, another mechanism of action besides nutrient benefit is indicated.
Final Report
Weeding Experiment
The weeding study was conducted in 2022 and 2023 at the University of Florida Plant Science Research and Education Unit (PSREU) near Citra, Florida. The field used in 2022 was a Candler sand (Hyperthermic, uncoated Lamellic Quartzipsamments) and the field used in 2023 was an Arredondo fine sand (Loamy, siliceous, semiactive, hyperthermic Grossarenic Paleudults). Weather was similar between years, with 1162 mm precipitation in 2022 and 1104 mm in 2023 (FAWN, 2024). The experiment was a randomized complete block design (RCBD) with factorial treatments. The first factor, cropping system, had 9 levels: sole cassava, sole Bambara nut, sole cowpea, sole peanut, sole soybean, cassava-Bambara intercrop, cassava-cowpea intercrop, cassava-peanut intercrop, and cassava-soybean intercrop. The second factor, weeding, had 2 levels: no weeding and full weeding. These factors combined for a total of 18 treatments replicated in 4 blocks.
An incorrect herbicide application in May 2022 necessitated the replanting of the weeding experiment, and due to lack of cassava planting material, the replanted 2022 experiment consisted only of the sole legume plots (4 out of the 9 levels of the cropping system factor); the 2023 iteration of the experiment included all cropping systems. The cassava used was Panama Sweet, an erect sweet variety (ECHO, Fort Myers, FL); cuttings approximately 10 cm long were planted vertically in the soil. The cowpea variety used was Red Ripper in 2022 and Iron Clay in 2023. The peanut variety used was H&W 136 in 2022 and Olin in 2023. E103 Bambara groundnut and Osage soybean were used in both 2022 and 2023. Legume seeds were treated with Exceed legume inoculant (Visjon Biologics, Henrietta, TX) to ensure adequate nodulation.
All plots were planted on June 29 in 2022 and on May 25 in 2023. Cassava was planted with a 1x1 m spacing for a total of 16 plants per plot and a density of 10,000 plants/ha. All legumes were planted at a 0.5x0.2 m spacing for a density of 100,000 plants/ha. Intercrop spacing was purely additive, with two rows of legumes between each two rows of cassava (0.25 m between cassava and legume rows). Plots were not fertilized but irrigated as needed. In 2022, full weeding consisted of manual weeding at 3, 6, and 9 weeks after planting; in 2023, full weeding consisted of Liberty (glufosinate-ammonium [2-amino-4-hydroxymethylphosphinyl)butanoic acid], BASF, Research Triangle Park, NC) herbicide application using a hooded sprayer at 4 and 8 weeks after planting.
Legume height was measured at maturity. Cowpea was harvested on September 26 in 2022 and on December 6 in 2023 (cowpea in 2023 had robust vegetative growth but did not produce flowers and pods until November). Soybean was harvested on October 5 in 2022 and on November 14 in 2023. Peanut was harvested on October 19 in 2022 and September 20 in 2023. Bambara groundnut suffered crop failure in both years. All legume harvests were carried out by hand. At each harvest, legume pods and residue were weighed, and subsamples were removed and dried to calculate dry mass of pods and residue. Cassava was harvested by hand on December 6 in 2023. Cassava roots and aboveground biomass were weighed, and subsamples (1 representative plant per plot) taken and dried to calculate dry mass. At every harvest, weeds were collected from a representative 90 cm2 quadrat in each harvested plot; samples were dried to determine weed dry biomass. Drying was performed in a forced-air oven at 60C for 72 hours or until dry weight had stabilized.
Data was organized in Microsoft Excel and analyzed using the lmerTest package in R, in linear mixed models with legume species, weeding regime, and year as fixed effects and block as a random effect. Figures were created using JMP. Land equivalent ratio (LER) was calculated from sole and intercropped cassava and legume yields.
Competition Pots
To further investigate competition between cassava and legumes, “competition pots” were created with artificial partitions to separate the above-ground and below-ground components of competition. Competition pot experiments were carried out in circular pots 30 cm deep and 60 cm in diameter (area of soil surface = 0.283 m2). Pots were filled by shovel with an Arredondo fine sand. Each pot was divided in half by a vertical barrier above the soil surface, below the soil surface, both above and below the soil surface, or neither (i.e. no barriers). Above-ground barriers consisted of hardware cloth with a 1 cm aperture. Below-ground barriers consisted of plywood sealed with caulk. Each pot was also encircled by a cylinder of hardware cloth 90 cm tall to prevent plants from growing beyond the 0.283 m2 area of the competition pot.
Competition pot experiments used a factorial randomized complete block design (RCBD), where the first factor was the presence or absence of above-ground separation and the second factor was the presence or absence of below-ground separation, for a total of 4 treatments replicated in 5 blocks. In each pot, a single cassava plant was grown in one half of the pot and 5 legume plants were grown in the other half, 25 cm from the cassava in a row with 7.5 cm between plants. The goal was to roughly approximate the spacing and density of a cassava-legume intercrop in the field. Competition pots were not fertilized but watered daily and hand-weeded throughout the experiment. The competition pot experiment was run twice, once with peanut and once with cowpea. The experiments were harvested on peanut or cowpea maturity. At harvest, legume height, biomass, and yield were measured, as well as cassava height and biomass. Competition pots give insight into how legumes are affected by competition with cassava and the condition of immature cassava at the time of harvest of intercropped legumes, ~3 months after planting.
Residue Experiment
This study was conducted in 2022 and 2023 at the University of Florida Plant Science Research and Education Unit (PSREU) near Citra, Florida. The field used in 2022 was a Gainesville loamy sand and the field used in 2023 was an Arredondo sand. Weather was similar between years, with 732 mm precipitation during the growing season in 2022 and 767 mm in 2023 (FAWN, 2024). The experiment was a randomized complete block design (RCBD) with factorial treatments. The first factor, cropping system, had 9 levels: sole cassava, sole Bambara nut, sole cowpea, sole peanut, sole soybean, cassava-Bambara intercrop, cassava-cowpea intercrop, cassava-peanut intercrop, and cassava-soybean intercrop. The second factor, residue management, had 2 levels: legume residues retained and legume residues removed. These factors combined for a total of 18 treatments replicated in 4 blocks. The cassava used was Panama Sweet, an erect sweet variety (ECHO, Fort Myers, FL). In 2022, rapid multiplication techniques were used to create small rooted shoot cuttings, approximately 10 cm tall, which were transplanted into the field. In 2023, woody cuttings approximately 10 cm long were planted vertically, directly in the soil. The cowpea variety used was Red Ripper in 2022 and Iron Clay in 2023. Olin peanut (a Spanish market type), E103 Bambara groundnut and Osage soybean were used in both 2022 and 2023. Legume seeds were treated with Exceed legume inoculant (Visjon Biologics, Henrietta, TX) to promote nodulation.
All plots were planted on May 5 in both 2022 and 2023. Cassava was planted with a 1x1 m spacing for a total of 16 plants per plot and a density of 10,000 plants/ha. All legumes were planted at a 0.5x0.2 m spacing for a density of 100,000 plants/ha. Intercrop spacing was purely additive, with two rows of legumes between each two rows of cassava. Plots were not fertilized and irrigated as needed. Cowpea was harvested on July 19 in 2022 and on October 20 in 2023 (cowpea in 2023 had robust vegetative growth but did not produce flowers and pods until October). Peanut was harvested on August 29 in both 2022 and 2023. Soybean suffered crop failure in 2022 and was harvested on October 13 in 2023. Bambara groundnut suffered crop failure in both years. All legume harvests were carried out by hand. At each harvest, legume pods and residue were weighed, and subsamples were removed and dried to calculate dry mass of pods and residue. Legume residues were either removed from plots or left on the soil surface, according to the residue management factor. To determine the impact of legume residues on sole cassava, residues removed from peanut plots were applied to those sole cassava plots which had the “residue retained” treatment. Litterbags were used in 2022 to track the decomposition of cowpea and peanut residues after harvest (Bocock and Gilbert, 1957). Labeled polypropylene mesh bags (~5 mm mesh) were filled with dried legume residues, weighed, and placed on the soil surface in corresponding plots. Three litterbags were placed in each plot after legume harvest, and individual litterbags were removed at intervals, ending with cassava harvest (cowpea litterbags were removed August 23, September 26, and November 21; peanut litterbags were removed September 26, October 19, and November 21). Removed litterbags were dried and weighed, and residue decomposition was calculated from the initial and final dry biomass in each bag (Bocock and Gilbert, 1957). Cassava was harvested by hand on November 21 in 2022 and on December 5 in 2023. Cassava roots and aboveground biomass were weighed and subsamples were removed and dried to calculate dry mass. Legume height was measured at maturity and cassava height was measured at harvest.
Data was organized in Microsoft Excel and analyzed using the lmerTest package in R, in linear mixed models with cropping system, residue management, and year as fixed effects and block as a random effect. Figures were created using JMP. Land equivalent ratio (LER) was calculated from sole and intercropped cassava and legume yields, and since LER is a response ratio, the natural logarithm of LER was used for analysis (Hedges et al., 1999).
Weeding Experiment
Legume species and weeding treatment both had significant effects on weed biomass at harvest and a significant interaction—plots that received full weeding had less weed biomass than unweeded plots, but this effect was most pronounced in soybean and sole cassava plots (Figure 1). Cowpea and peanut plots had low weed biomass regardless of weeding treatment, comparable to fully weeded soybean or sole cassava plots. The ability of cowpea and peanut to suppress weed growth may be due in part to their greater height: at 8 weeks after planting, cowpea, peanut, and soybean plants had an average height of 64.6 cm, 41.4 cm, and 36.5 cm, respectively, all higher than the average cassava height of 21.2 cm. Linear regression of weed biomass and legume height showed that weed biomass decreases as legume height increases, and that this effect is stronger in unweeded plots than in weeded plots (Figure 2).
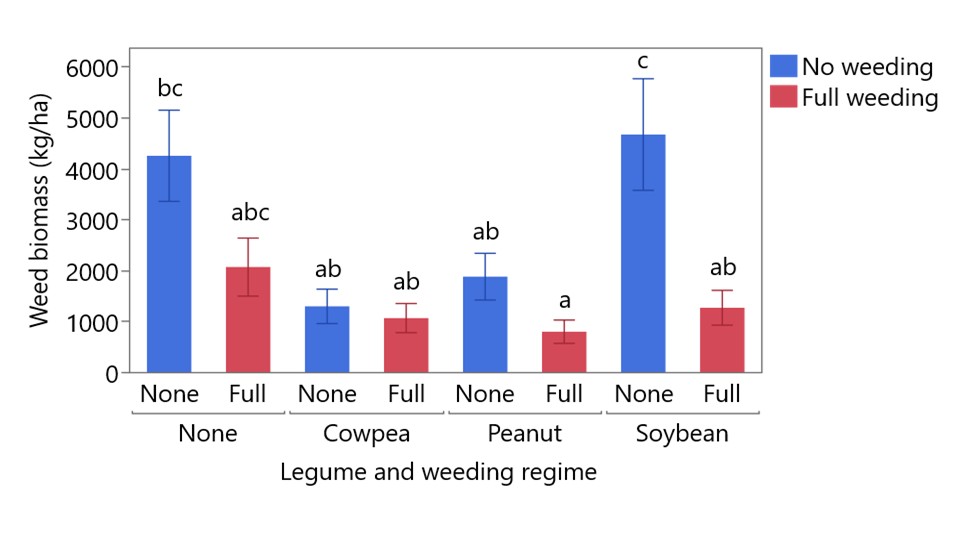
Figure 1. Effect of legume and weeding regime on weed biomass at harvest. Error bars are standard error, bars sharing the same letter are not significantly different (p > 0.05).
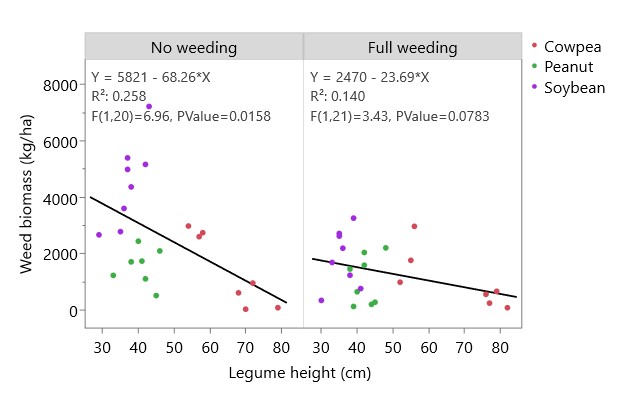
Figure 2. Effect of legume height and weeding regime on weed biomass at harvest.
Legume yield varied by year: in 2022 pod yield of cowpea (489 kg/ha) was higher than peanut (107 kg/ha) and soybean (169 kg/ha), whereas in 2023 pod yield of peanut (631 kg/ha) and soybean (596 kg/ha) were higher than cowpea (217 kg/ha) (Figure 3). Total crop biomass in each system (including intercropping with cassava) followed a similar pattern (Figure 4). Crop biomass proportion (crop biomass/weed biomass + crop biomass) was calculated to reflect the efficiency of each legume and weeding regime combination in increasing the proportion of plant biomass not found in weeds. Sole cassava (regardless of weeding regime) and unweeded soybean had significantly lower crop biomass proportions than all other combinations, with less than 20% of biomass as crops in the former group and more than 50% of biomass as crops in the latter group (Figure 5). Fully weeded peanut plots had the highest crop biomass proportion at 0.76, i.e. 76% of biomass as crops and 24% as weeds. Cassava yield was higher in weeded plots (627 kg/ha) than in unweeded plots (209 kg/ha), but this difference was not statistically significant (p = 0.108). Yields of sole and intercropped cassava and legumes in each weeding regime are shown in Table 1, along with their associated partial LERs and the total LER for each system. Average LER was 2.57 for weeded systems and 3.56 for unweeded systems.
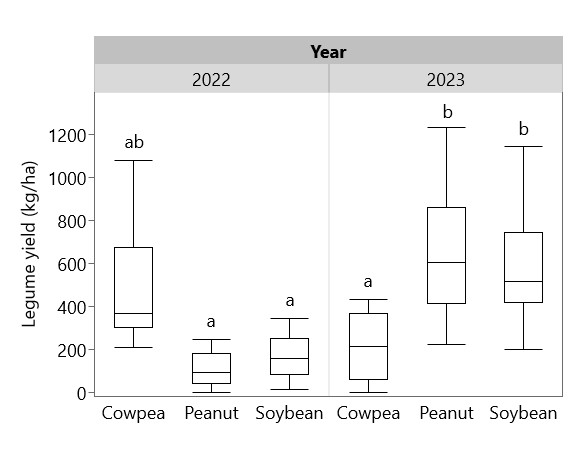
Figure 3. Legume yields in 2022 and 2023. Box plots sharing the same letter do not have significantly different means (p > 0.05)
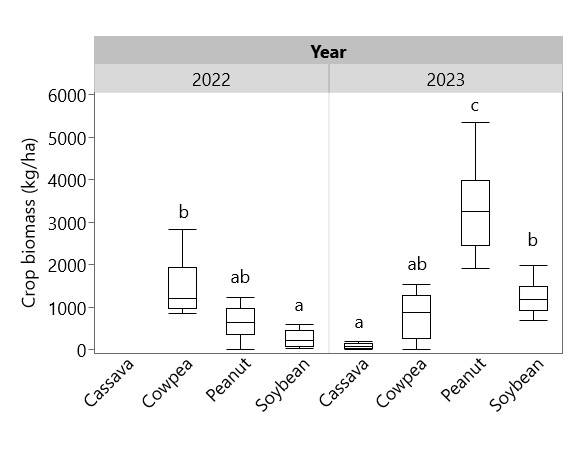
Figure 4. Total crop biomass in 2022 and 2023. Box plots sharing the same letter do not have significantly different means (p > 0.05).
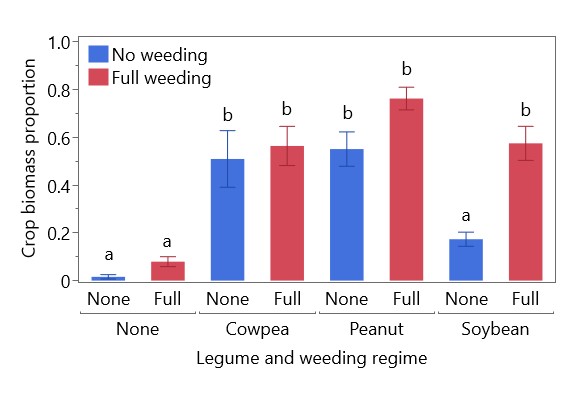
Figure 5. Effect of legume and weeding regime on crop biomass proportion (total crop biomass divided by total plant biomass, including weeds). Error bars are standard error, bars sharing the same letter are not significantly different (p > 0.05).
Table 1. LER of weeded and unweeded cassava-legume intercropping systems in 2023. LER is the sum of partial LER (PLER) of each component crop.
Legume yield (kg/ha) Cassava yield (kg/ha)
| System | Sole | Intercroppped | PLER | Sole | Intercropped | PLER | LER |
| Full Weeding | |||||||
| Cowpea | 280 | 174 | 0.62 | 383 | 808 | 2.11 | 2.73 |
| Peanut | 490 | 575 | 1.17 | 383 | 78 | 0.20 | 1.38 |
| Soybean | 732 | 599 | 0.82 | 383 | 1403 | 3.67 | 4.49 |
| No weeding | |||||||
| Cowpea | 197 | 220 | 1.11 | 104 | 227 | 2.18 | 3.30 |
| Peanut | 607 | 852 | 1.40 | 104 | 227 | 2.18 | 3.59 |
| Soybean | 512 | 542 | 1.06 | 104 | 287 | 2.76 | 3.82 |
The results of the weeding experiment support the use of intercropped legumes as a weed management technique in cassava-legume intercropping. The capacity of legumes to suppress weeds was correlated with their vigor and height: plots with cowpea and peanut, which were taller than soybean and qualitatively more vigorous, had less weed biomass than plots with soybean. Unweeded soybean had the highest weed biomass, and soybean correspondingly received a greater benefit from weeding than cowpea and peanut plots.
The slow early growth of cassava makes it particularly susceptible to weed pressure, and weed biomass was high in sole cassava plots, even when the full weeding treatment was applied. Intercropping cowpea and peanut reduced weed biomass to an extent comparable to full weeding, and the combination of peanut and full weeding produced the lowest weed biomass. Legume and cassava yields were not significantly affected by intercropping, suggesting that the use of intercropped legumes for weed management does not have an undue yield penalty. On the contrary, high LER values in these systems indicated a yield benefit of cassava-legume intercropping, particularly in the unweeded treatments (average LER = 3.56 vs 2.57 in weeded treatments). This result supports the Stress Gradient Hypothesis (SGH), which predicts that the removal of a biotic stress (e.g. weed pressure, removed by weeding) shifts plant interactions toward competition (i.e. lower LER). These results also suggest that the yield benefit of cassava-legume intercropping is due in part to the ability of intercropped legumes to suppress weeds in the first few months after planting.
Competition Pots
Analysis of cassava and legume height, yield, and biomass did not reveal significant interactions between the effects of above-ground separation and below-ground separation. Table 2 shows the main effects of above and below-ground separation, and all the statistically significant effects were caused by above-ground competition. In cassava-peanut competition pots, peanut height was significantly lower in the presence of above-ground competition (Table 2). Peanut biomass, cassava height, and cassava biomass were also lower in the presence of above-ground competition, but these effects were not significant.
Table 2. Effect of above-ground and below-ground competition on peanut and cassava. P values calculated from one-way ANOVA. Cowpea height listed as 60 cm because all cowpea plants reached the maximum height of the competition plots.
| System | Isolated | Above-ground competing | p | Isolated | Bel0w-ground Competing | p |
| Peanut-cassava | ||||||
| Peanut height (cm) | 33.8 | 30.3 | 0.031 | 32.1 | 32.7 | 0.731 |
| Cassava height (cm) | 34.6 | 26.2 | 0.062 | 30.9 | 31.6 | 0.883 |
| Peanut biomass (g) | 138.1 | 102.8 | 0.062 | 30.9 | 116.8 | 0.493 |
| Cassava biomass (g) | 16.3 | 13.1 | 0.125 | 14.5 | 15.6 | 0.617 |
| Peanut yield (g) | 15.6 | 13.2 | 0.555 | 16.1 | 13.0 | 0.432 |
| Cowpea-cassava | ||||||
| Cowpea height (cm) | 60 | 60 | 60 | 60 | ||
| Cassava height (cm) | 37.0 | 33.6 | 0.484 | 36.7 | 33.1 | 0.459 |
| Cowpea biomass (g) | 88.3 | 143.9 | 0.069 | 126.3 | 115.1 | 0.725 |
| Cassava biomass (g) | 2.1 | 1.2 | 0.042 | 1.8 | 1.2 | 0.233 |
| Cowpea yield (g) | 34.8 | 56.7 | 0.069 | 49.8 | 45.3 | 0.725 |
Cowpea, unlike peanut and cassava, was more productive in the presence of above-ground competition, with higher biomass and yields (Table 2). Cassava biomass was significantly lower in the presence of above-ground competition (2.1 g isolated vs 1.2 g competing, p = 0.042). The cassava biomass proportion (cassava biomass/cowpea biomass + cassava biomass) was not affected by below-ground competition, but was significantly higher with above-ground separation, where cassava did not have to compete with cowpea for light (Figure 6). Above-ground competition with cowpea did not significantly affect cassava height, but was associated with more spindly cassava stems, with less weight per unit height (Figure 7).
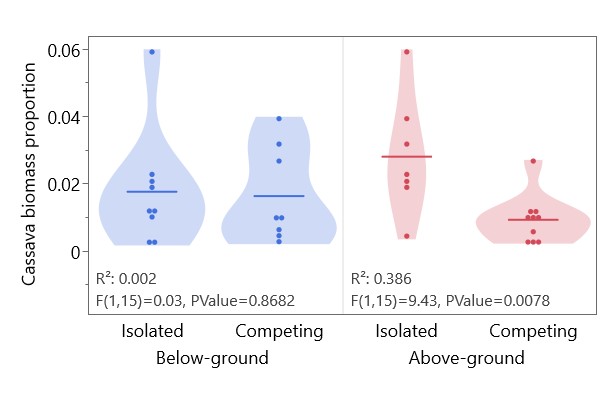
Figure 6. Effect of above-ground and below-ground competition on cassava biomass proportion (total cassava biomass/total pot biomass). Data from cassava-cowpea competition pots. Horizontal lines represent population means. No significant interaction between above-ground and below-ground effects.
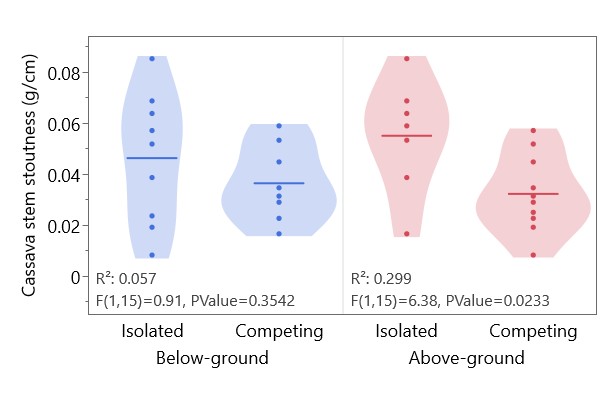
Figure 7. Effect of above-ground and below-ground competition on cassava stem stoutness (g dry biomass/cm stem length). Data from cassava-cowpea competition pots. Horizontal lines represent population means. No significant interaction between above-ground and below-ground effects.
Competition pot experiments showed that cowpea outcompetes cassava while peanut and cassava are more competitively balanced. Above-ground competition in the cassava-peanut system led to lower heights and biomass in both peanut and cassava. Above-ground competition in the cassava-cowpea system, on the other hand, led to lower cassava biomass but higher cowpea biomass and yield. Competition with cowpea also induced a spindly phenotype in cassava; cassava plants competing above-ground with cowpea had significantly less biomass per unit height, likely in an effort to grow beyond the dense cowpea canopy (Craine and Dybzinski, 2013). The competitive vigor of cowpea must be managed to prevent undue reduction of cassava growth in cassava-cowpea intercropping systems; for example, planting cassava earlier than cowpea may allow it to reach a more competitive height before cowpea canopy closure, and lower cowpea density or less vigorous cowpea varieties can be used to the same effect (Cenpukdee and Fukai, 1992a).
In both cassava-cowpea and cassava-peanut competition pots, above-ground competition was more impactful than below-ground competition. This suggests that competition for light, more than competition for water or nutrients, drives interactions between cassava and legumes in this environment (the effect of environment should not be ignored; this experiment was irrigated daily, making competition for water unlikely) (McPhee and Aarssen, 2001). Observation of the competition pots supports this hypothesis: in pots with above-ground separation, plants from both sides grew up against the hardware cloth barriers, and in pots without above-ground separation, the cassava and legume canopies mingled, mostly due to cowpea and peanut leaves and stems growing into the cassava halves of the pots. Cassava and peanut were similar in height while cowpea grew much taller, accessing a greater portion of light and significantly reducing cassava biomass when given the opportunity to overshadow cassava.
Residue Experiment
Legumes were generally unaffected by intercropping in this experiment. Legume yields were significantly higher in 2022 than in 2023, but there was no significant difference in yields between sole legumes and intercropped legumes (Figure 8). Legume heights at maturity were significantly different between species (p < 0.0001), with cowpea the tallest (71.3 cm), followed by peanut (53.4 cm) and soybean (38.6). Decomposition of legume residues on the soil surface after legume harvest is shown in Figure 9. Decomposition was most rapid in the first 30 days after harvest, reaching ~40% of initial biomass, then continued gradually to 50.3% of peanut residues and 62.5% of cowpea residues at cassava harvest. Based on these decomposition rates, an estimated 3864 kg/ha of cowpea residue and 6442 kg/ha of peanut residue had decomposed in residue-retained plots by the time of cassava harvest in 2022, and 2027 kg/ha cowpea residue and 785 kg/ha peanut residue decomposed by cassava harvest in 2023. Using previously determined rates of N content in legume residues (Kermah et al., 2018), these decomposed biomass quantities can be estimated to contain 98 kg N/ha, 189 kg N/ha, 52 kg N/ha, and 23 kg N/ha, respectively.
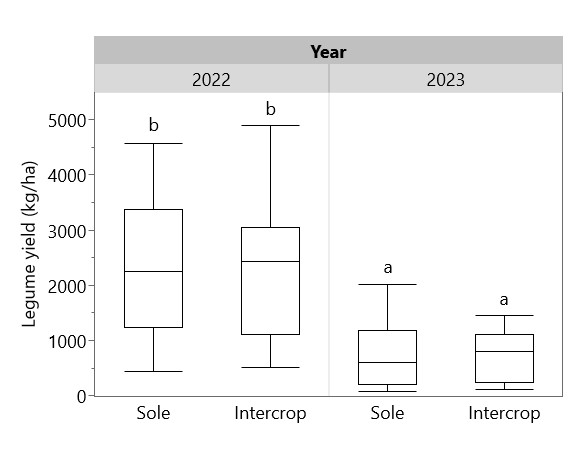
Figure 8. Sole and intercropped legume yields in 2022 and 2023. Box plots include yields of all legumes (n = 24 for each year/system combination). Box plots sharing the same letter do not have significantly different means (p > 0.05).
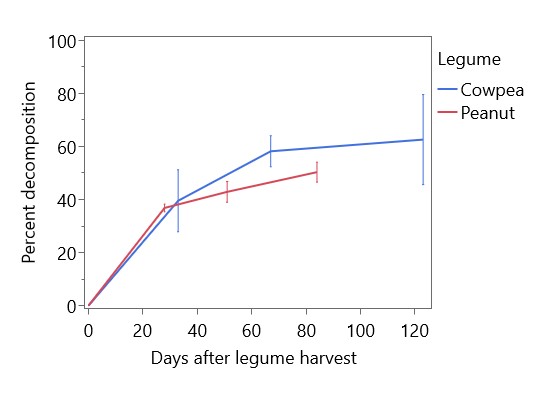
Figure 9. Decomposition of cowpea and peanut residues determined by litterbags. Final timepoints (84 days after harvest for peanut and 123 days after harvest for cowpea) were taken at cassava harvest. Error bars represent 95% confidence intervals.
Cassava height at harvest was greater in 2022 (119.3 cm) than in 2023 (66.1 cm), and intercropped legume species had a significant effect on cassava height in 2022, with cowpea suppressing cassava height (98.4 cm) compared to peanut (120.3 cm), soybean (124.6 cm) and sole cassava (129.3 cm) (Figure 10). Intercropping with cowpea was also associated with higher cassava mortality—only 60% of cassava plants intercropped with cowpea survived to harvest, compared with 82% with peanut, 90% with soybean, and 91% with sole cassava (Figure 11). Cassava yield in 2022 ranged from 2866 kg/ha intercropped with cowpea to 10321 as a sole crop, whereas in 2023, cassava yields were consistently low, ranging from 1233 kg/ha intercropped with soybean to 1658 kg/ha with peanut (Figure 12). Figure 13 shows that while intercropped cassava yielded significantly less than sole cassava in 2022, intercropping produced no difference in cassava yield in 2023. Residue management did not significantly affect cassava yield.
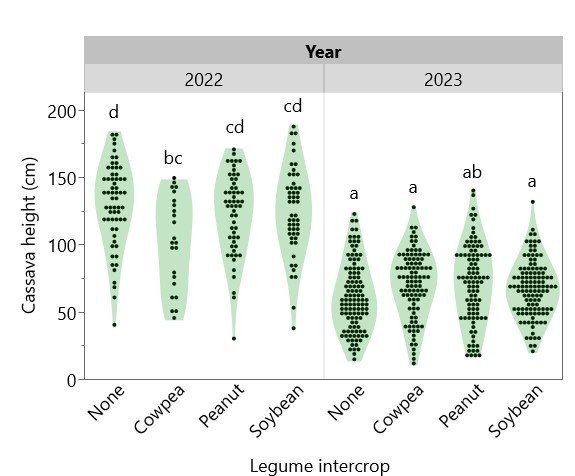
Figure 10. Effect of legume intercrop on cassava height in 2022 and 2023. Legume intercrop “None” indicates sole cassava. Each point represents one cassava plant at harvest. Violin plots sharing the same letter do not have significantly different means (p > 0.05).
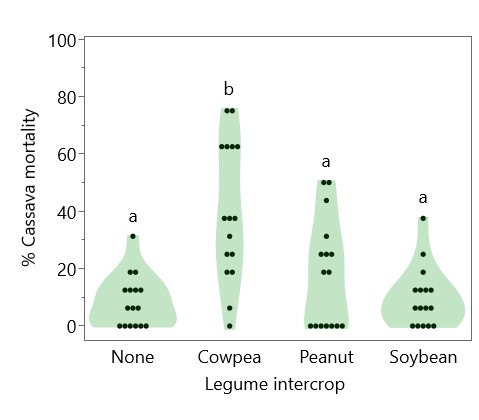
Figure 11. Effect of legume intercrop on cassava mortality. Legume intercrop “None” indicates sole cassava. Each point represents one plot. Violin plots sharing the same letter do not have significantly different means (p > 0.05).
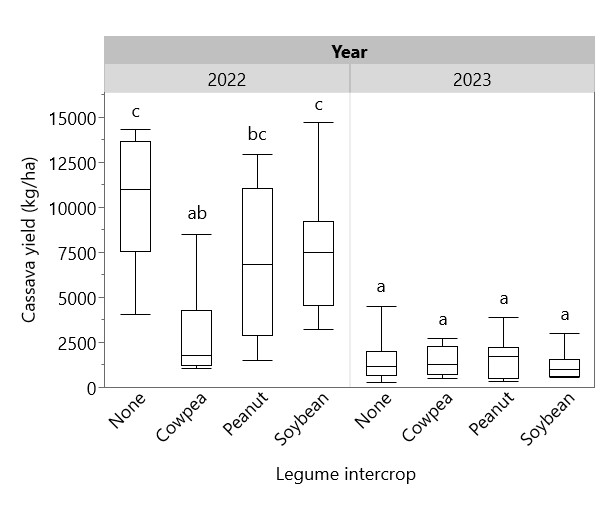
Figure 12. Effect of legume intercrop on cassava yield in 2022 and 2023. Legume intercrop “None” indicates sole cassava. Box plots sharing the same letter do not have significantly different means (p > 0.05).
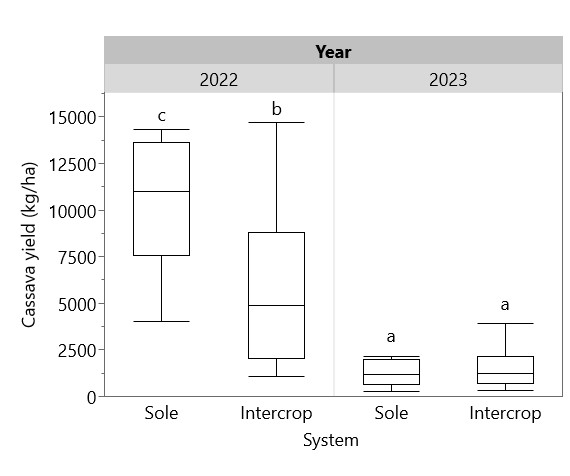
Figure 13. Sole and intercropped cassava yields in 2022 and 2023. Box plots sharing the same letter do not have significantly different means (p > 0.05).
Yields of sole and intercropped cassava and legumes are shown in Table 3, along with the corresponding partial LERs and overall LER of each system in each year. LER was > 1 in all systems, indicating a yield benefit of cassava-legume intercropping that ranged from mild (with cowpea in 2022, LER = 1.21) to dramatic (with peanut in 2023, LER = 2.71) (Figure 14). Residue management did not significantly affect overall LER, but cassava partial LER in 2023 was significantly higher with residues removed (Figure 15). To test the prediction of the Stress Gradient Hypothesis that plant interactions shift towards facilitation in more stressful environments, linear regressions of LER with sole cassava yield and sole legume yield were conducted and found significant negative relationships between sole yield and LER (Figure 16). Similar negative correlations were found between LER and sole cassava residue, sole legume residue, and overall sole crop biomass (Figure 17). There was no significant relationship between LER and intercropped crop yields or biomass.
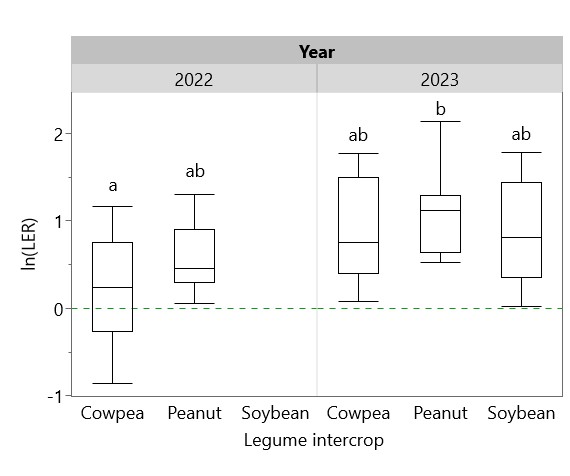
Figure 14. Effect of legume intercrop on cassava-legume LER in 2022 and 2023. Dotted green line at ln(LER) = 0 indicates additivity; ln(LER) > 0 indicates a yield benefit of intercropping. Box plots sharing the same letter do not have significantly different means (p > 0.05).
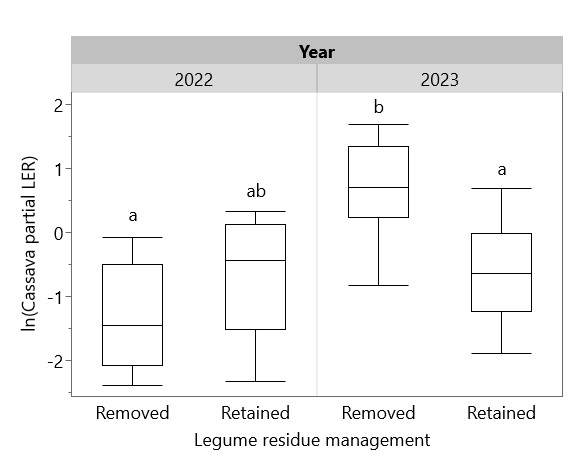
Figure 15. Effect of legume residue management on cassava partial LER in 2022 and 2023. Cassava partial LER calculated as intercropped cassava yield/sole cassava yield. Box plots sharing the same letter do not have significantly different means (p > 0.05).
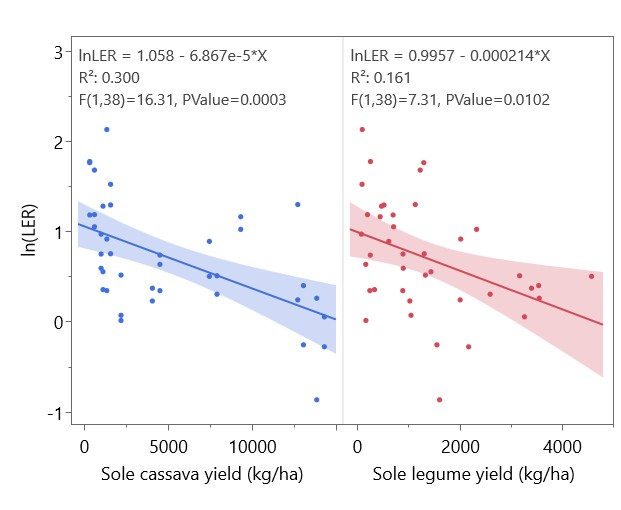
Figure 16. Linear regression of LER with sole cassava and legume yields. Shaded areas represent 95% confidence regions for linear fit. Intercropping has a yield benefit where ln(LER) > 0.
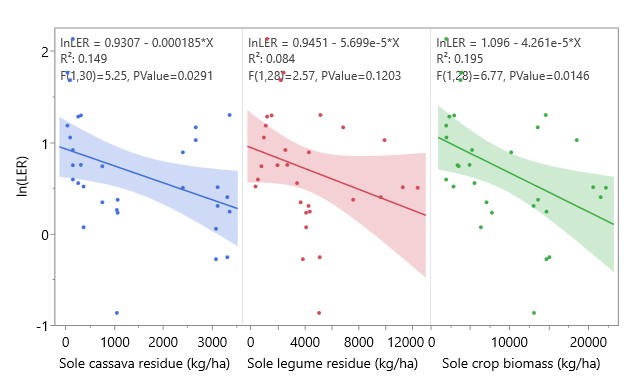
Figure 17. Linear regression of LER with sole cassava and legume residue production and total sole crop biomass. Residue was defined as dry mass of leaves, stems, and other non-yield plant parts. Total sole crop biomass was defined as cumulative biomass of sole cassava and legume plots. Shaded areas represent 95% confidence regions for linear fit. Intercropping has a yield benefit where ln(LER) > 0.
Table 3. LER of cassava-legume intercropping systems by legume intercrop and legume residue management in 2022 and 2023. LER is the sum of partial LER (PLER) of each component crop.
Legume yield (kg/ha) Cassava yield (kg/ha)
| Residue | Sole | Intercropped | PLER | Sole | Intercropped | PLER | LER |
| 2022 | |||||||
| Removed | |||||||
| Cowpea | 2076 | 1784 | 0.86 | 11989 | 2512 | 0.21 | 1.07 |
| Peanut | 2771 | 3405 | 1.23 | 11989 | 5661 | 0.47 | 1.70 |
| Retained | |||||||
| Cowpea | 915 | 908 | 0.99 | 8654 | 3221 | 0.37 | 1.36 |
| Peanut | 3459 | 2746 | 0.79 | 8654 | 8144 | 0.94 | 1.73 |
|
|
|
|
|
|
|
|
|
| 2023 | |||||||
| Removed | |||||||
| Cowpea | 1456 | 1004 | 0.69 | 921 | 2140 | 2.32 | 3.01 |
| Peanut | 498 | 1032 | 2.07 | 921 | 1814 | 1.97 | 4.04 |
| Soybean | 189 | 239 | 1.26 | 921 | 1531 | 1.66 | 2.93 |
| Retained | |||||||
| Cowpea | 1059 | 1025 | 0.97 | 2169 | 794 | 0.37 | 1.33 |
| Peanut | 729 | 822 | 1.13 | 2169 | 1503 | 0.69 | 1.82 |
| Soybean | 178 | 179 | 1.01 | 2169 | 936 | 0.43 | 1.44 |
All cassava-legume systems in this experiment had LER > 1 in both 2022 and 2023, indicating a consistent yield benefit from cassava-legume intercropping. (Table 3) Legumes yields were not significantly affected by intercropping, suggesting that the legumes were highly complementary with cassava or that the legumes were competitively dominant over cassava. Data from cassava intercropped with cowpea and peanut in 2022 support the latter option: cassava yields were lower in intercropped than sole, and cassava intercropped with cowpea was significantly shorter and had a lower survival rate than sole cassava. In most cassava-legume intercropping systems, cassava is dominant (Leihner, 1983; Mutsaers et al., 1993). The dominance of legumes in this experiment (cowpea in particular) could be due to slow cassava growth rates in response to nutrient stress (El-Sharkawy, 2004), as the study was carried out on sandy soil with no inputs of fertilizer. Cowpea had the greatest negative effect on cassava growth and yield, possibly due to cowpea being the tallest legume, and qualitatively the most vigorous.
Litterbag data showed that cowpea and peanut residues decomposed quickly enough for roughly half of residue nutrients to enter the soil in the time between legume harvest and cassava harvest. There was no major difference in decomposition rates between cowpea and peanut residues, but since cowpea was harvested earlier in the season, nutrients from cowpea residues may have been available to cassava sooner. Nutrients from decomposing residues are generally immobilized by soil microbes for a time depending in part on the C:N ratio of the residues, so these nutrients are not immediately available to plants. A previous study of cowpea and peanut residues in the context of cassava intercropping found that cowpea and peanut residue nutrients were immobilized for 30-70 days and 30-45 days after application, respectively, after which net mineralization occurred; however, this data was from an incubation of legume residues mixed with soil, not in field conditions (Gnahoua et al., 2016). Decomposition of legume residues can be sped up by incorporation of the residues into the soil (Sanchez, 2019), but this is not practical in cassava-legume intercropping as the digging required for residue incorporation would likely damage cassava storage roots. In addition to potential nutrient benefits from decomposition, legume residues may also affect cassava by decreasing soil evaporation, harboring microbes and insects, and lowering soil temperature.
Removal of residues did not directly affect cassava yield but had a significant positive impact in 2023 on cassava partial LER—the ratio of intercropped cassava yield to sole cassava yield. Correspondingly, retaining legume residues increased yields of sole cassava more than it increased yields of intercropped cassava. This could be because sole cassava plots, which contained no legume biomass prior to the addition of peanut residues, were limited by nutrient availability, while intercropped cassava plots were limited by competition with legumes. However, the impacts of legume residue removal and retention are inconsistent between 2022 and 2023, and further study is needed to unpick the costs and benefits of legume residue retention in cassava-legume intercropping. For the purpose of farmer decision-making, the yield benefit of retaining legume residues must be balanced against the benefit of removing residues for use as animal feed or for sale; the labor cost of residue removal must also be taken into account.
Educational & Outreach Activities
Participation summary:
N/A
Project Outcomes
Summary and Conclusions
The studies found that cassava-legume intercropping is generally beneficial in terms of yield. Cowpea, peanut, and soybean all exhibited Land Equivalency Ratio > 1 in combination with cassava. Of the legumes studied, peanuts were found to be the best intercrop with cassava, exhibiting weed-suppressing abilities comparable to cowpea, but not outcompeting cassava as severely. The weeding experiment found that tall, vigorous legumes can suppress weeds to an extent comparable to manual weeding in the first two months after planting. An additive effect of weeding and legume intercropping was seen in the system with lowest weed biomass (weeded peanut). The competition pot experiment showed that above-ground competition plays a more important role in cassava-legume systems than below-ground competition, a result supported by the importance of legume height observed in the weeding experiment and in the residue experiment. Correspondingly, the competition pot experiment also showed that cowpea competes more heavily with cassava than does peanut, suggesting that cassava-peanut systems may be easier to balance competitively than cassava-cowpea systems. In the residue experiment, cassava had higher mortality, lower height, and lower yield when intercropped with cowpea than in any other system; cassava-peanut had the highest LER in each year and in each residue management treatment. The residue experiment did not detect a significant effect of legume residue management on cassava yield. However, cassava partial LER was higher in the residue removed treatment, possibly due to different limiting factors between sole cassava and intercropped cassava. Cowpea and peanut residues left on the soil surface decomposed significantly in the time between legume harvest and cassava harvest: roughly 40% of residue biomass decomposed in the first month after legume harvest, and a total of 50-60% had decomposed by the time of cassava harvest. Immobilization of nutrients or insufficient time for nutrients to affect yield formation could have contributed to the lack of observable effect of residue management on cassava yield.
For Florida, the main application is that cassava-legume intercropping is possible and exhibits a yield benefit, but cassava intercropping is not likely to be applied on any commercial scale in Florida. However, it may be of interest to the growing homesteading and backyard gardening movements throughout the state. For cassava-legume intercropping throughout the world, this research supports the use of short, short-duration grain legumes as intercrops with cassava that can effectively suppress weeds without undue damage to cassava yields. Scientific study of cassava-legume systems also represents a necessary step in the development of extension materials for cassava-legume intercropping, materials which are currently lacking at most scales. The lack of a significant cassava yield benefit from legume residue retention in this study suggests that farmers may find more advantage from using legume residues as livestock feed, though this question requires further study and will depend on many local factors.
N/A