Final report for GW21-223
Project Information
More frequent mega-wildfires reshaping Great Basin plant communities, endangering native perennial grasses that are a key source of forage for livestock. Predicted increase in the frequency of drought and ongoing invasion by cheatgrass (Bromus tectorum) will exacerbate the impact of wildfires in the Great Basin. Post-fire seeding has become a common practice to restore degraded lands to native perennial grasslands, but whether restored communities will persist in the future is unknown. Our research aims to understand how forward-looking post-fire seeding can promote resilient rangelands in the Great Basin, which can safeguard the economic viability of ranching and the quality of life of ranchers from catastrophic wildfires. We assess the capacity of current seeding practices and potential seed source modification to withstand future drought and invasion. First, we survey vegetation in the boundaries of a large fire to determine the effects of post-fire seeding and landscape dispersal on the species and genetic diversity of restored communities. Second, we use a common garden approach to understand how seed provenance and species traits alter the response of native grass populations to cheatgrass and drought. Our research will provide practical recommendations on where and what to plant in future post-fire restoration. We will disseminate results to ranchers and public land managers in Oregon and Idaho via field workshop, oral presentations, and newsletter articles. We expect increase in stakeholders’ knowledge about the effects of current post-fire seeding practices on plant diversity and the selective seed sourcing strategy to adapt to future drought and cheatgrass invasion.
Research objective 1: Assess the effects of current post-fire seeding practice on local plant species and genetic diversity in burned areas.
Research objective 2: Evaluate where on the landscape to prioritize post-fire seeding for native vegetation recovery in burned areas.
Research objective 3: Identify native grass seed provenance that improves the drought tolerance of restored sites.
Research objective 4: Understand the relationship between the drought tolerance and cheatgrass resistance traits of native grasses.
Education objective 1: Ranchers and land managers in the Northern Great Basin learn how current post-fire seeding practices affect the resilience of rangelands and how selective seed sourcing can aid in the adaptation to future drought and cheatgrass invasion.
Education objective 2: College and high school students in Oregon will learn about the importance of post-fire rangeland restoration.
Education objective 3: Range and restoration professionals will learn how the concept of pre-emptive restoration can be applied in post-fire seeding of arid rangelands.
Pre = before Sept 2021; Year 1 = Sept 2021 – Aug 2022; Year 2 = Sept 2022 – Aug 2023
|
Category |
Tasks |
Pre |
Year 1 |
Year 2 |
|
Research Objective 1 & 2 |
Field collection of leaves and plant composition Sequence DNA from leaf samples Analyze plant composition and genomic data Write Manuscript 1 |
X
|
X X |
X |
|
Research Objective 3 & 4 |
Field sampling of plant traits Perform carbon isotope analysis Analyze trait data Write Manuscript 2 |
X |
X X
|
X X |
|
Education Objective 1 |
Quarterly reports to the collaborating producer Lead post-fire restoration field workshop Oral presentations at Cattlemen’s Association meetings Oral presentation at a BLM field office Write short newsletter articles |
|
X |
X X X X X |
|
Education Objective 2 |
Rangeland restoration field class at the University of Oregon Recruit undergraduate students for research opportunities High school and college range camps |
|
X X X |
X X |
|
Education Objective 3 |
Oral presentation at the Society of Range Management meeting Poster presentation at the Society of Ecological Restoration Submit Manuscript 1 in Restoration Ecology Submit Manuscript 2 in Journal of Applied Ecology Website development Website and social media updates |
|
X X |
X X X X
X |
Here we highlight the milestones we plan on achieving between September 2021 and August 2023:
Research Objective 1 & 2: Aoyama will finish sequencing leaf samples by February 2022. Aoyama will finish analyzing plant community and genetic data by May 2022 and write a full draft of Manuscript 1 by September 2022. Research Objective 3 & 4: Aoyama will finish collecting trait data by July 2022, and finish carbon isotope analysis by August 2022. Aoyama will finish analyzing trait data by November 2022 and write a full draft of Manuscript 2 by February 2023.
Education Objective 1: Aoyama will report back to Mr. Hoagland, the collaborating producer, quarterly. The team will develop the field workshop material by March 2023, publicize the event in April-May, and host the event on June 2023. Mr. Hoagland and Aoyama will design and print out the brochures by February 2023 and give talks at the Oregon and Idaho Cattlemen’s Association meetings and at the BLM Owyhee and Burns field offices in spring 2023. Education Objective 2: Aoyama will co-instruct a field ecological methods course at the University of Oregon in winter quarter of 2022. Aoyama will recruit and mentor at least two undergraduate students between September 2021 and August 2023. Aoyama will lead a native plant restoration module in range camps in June 2022 and June 2023. Education Objective 3: Aoyama will give talks at the Society of Range Management annual meeting in February 2023 and the Society of Ecological Restoration conference in July 2023. The team will submit Manuscript 1 by November 2022 and Manuscript 2 by March 2023. Aoyama will set up a website by October 2021 and keep our research and outreach pages up to date.
Cooperators
- - Producer
Research
Objective 1. Assess the effects of current post-fire seeding practice on local plant species and genetic diversity in burned areas. We predict that native grass seeding will increase local species and genetic diversity in burned-seeded areas relative to burned-unseeded areas, because burned-unseeded areas are dominated by cheatgrass. We focus on plant diversity at species and genetic level here because we are concerned about the adaptive capacity of plant communities after post-fire seeding. Based on the resilience framework, species diversity buffers against multiple stressors such as drought (Ives and Carpenter 2007, Hautier et al. 2014), and genetic diversity ensures the populations’ potential to evolve when conditions change in the future (Hoffmann and Hercus 2000). Our hypothesis will be supported if species diversity and genetic diversity in burned-seeded areas are higher than those in burned-unseeded areas. Alternatively, seeded areas may have lower diversity than unseeded areas, due to low diversity in seed mixes which contain only 4-5 species from 1-2 source populations (Leger and Baughman 2015). The effect of these low diversity seed mixes on plant diversity of restored communities is not well tested but is likely to reduce the resilience of native perennial grasslands.
Objective 2. Evaluate where on the landscape to prioritize post-fire seeding for native vegetation recovery in burned areas. We predict that dispersal will increase recovery of native species and genetic diversity closer to fire edges, making post-fire seeding more important farther away from the fire edges. Wind dispersal of seeds from unburned areas is the natural process of post-fire recovery in burned areas (Damschen et al. 2014). Seed dispersal exponentially decreases with distance to the nearest seed source or fire edge (Nathan and Muller-Landau 2000). Therefore, in large wildfires, seeds may not reach some burned areas because the distance between unburned areas may be too great for the seeds to disperse. Post-fire seeding attempts to overcome this seed limitation (Foster and Tilman 2003). The effect of seeding on plant diversity may depend on distance from the nearest fire edge. We expect post-fire seeding in the core of these large burns will be most helpful for maintaining a resilient landscape. Our hypothesis will be supported if the effect size of seeding (difference in species and genetic diversity between seeded and unseeded areas) increases with distance from the nearest fire edge.
Objective 3. Identify native grass seed provenance that improves the drought tolerance of restored sites. We predict that the southern populations of native grass species will exhibit higher drought tolerance than the northern populations. This drought tolerance will help provide reliable forage in a hotter/drier future. Our preliminary results indicate that seeds sourced from southern populations in drier sites have higher seedling success rate when grown in a common garden than seeds from local or northern populations in wetter sites (Fig. 1).  We expect that this effect is associated with more conservative traits (such as higher water use efficiency) in these populations. A plant trait-based approach using simple-to-measure aboveground traits has been successful in explaining trait-by-environment associations across species (e.g., Baughman et al. 2019, Basey et al. 2015). We will test the association between seed provenance, plant traits, and drought survival. Our hypothesis will be confirmed if southern populations have higher water use efficiency, lower water stress, and higher survival rates than northern populations under drought.
We expect that this effect is associated with more conservative traits (such as higher water use efficiency) in these populations. A plant trait-based approach using simple-to-measure aboveground traits has been successful in explaining trait-by-environment associations across species (e.g., Baughman et al. 2019, Basey et al. 2015). We will test the association between seed provenance, plant traits, and drought survival. Our hypothesis will be confirmed if southern populations have higher water use efficiency, lower water stress, and higher survival rates than northern populations under drought.
Objective 4. Understand the relationship between the drought tolerance and cheatgrass resistance traits of native grasses. Keeping cheatgrass at bay is important because we expect a tradeoff between drought-tolerant and cheatgrass-resistant traits will decrease survivorship of native grasses under combined drought and invasion. Drought-tolerant individuals have resource-conservative traits (e.g., high water use efficiency (Farquhar et al. 1989) and low growth rates (Craine et al. 2012)) to save energy. In contrast, cheatgrass-resistant individuals are known to have resource-acquisitive traits (e.g., early phenology and high growth rates (Leger et al. 2019)) to compete with annual grasses that emerge in winter and senesce by summer. However, the competitive outcomes for restored natives vs. cheatgrass under contrasting precipitation conditions are still unknown. Understanding the trait association with seedling performance under drought and invasion would inform practitioners which native species and population to use for pre-emptive seeding practices depending on the extent of cheatgrass invasion or the likelihood of future drought in the area. Our hypothesis will be supported if native seedling survival rate increases with drought-tolerance traits under drought but decreases under drought and invasion, and vice versa.
RESEARCH METHODS AND MATERIALS. We tested our hypotheses using two approaches: (1) a Survey where we measure plant diversity at a community and population level in a previously burned area; and 2) a Common Garden Experiment where we measure intraspecific variability in native grass response to drought and cheatgrass treatments.
Objective 1 & 2: Survey of post-fire plant communities.
Sampling design. Our survey took place in Soda Fire, where over 280,000 acres of rangelands burned at the border of Oregon and Idaho in 2015 (MTBS; Fig. 2). 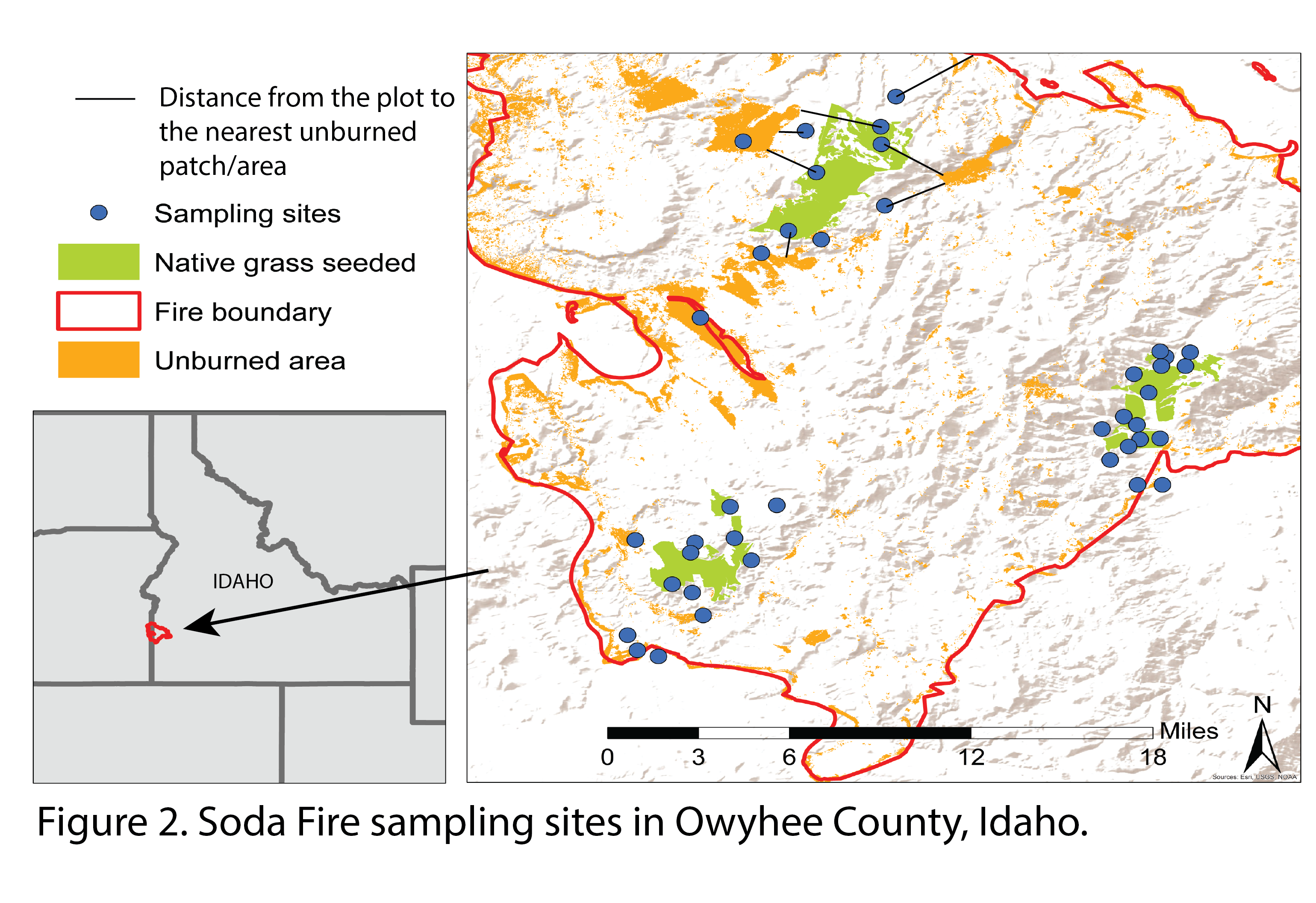 Taking advantage of existing USGS monitoring effort, we identified 40 plots stratified by burned-unseeded (n = 15), burned-seeded (n = 15), and unburned-unseeded (n = 10) areas. Using distance from the nearest fire edge as a proxy for landscape dispersal, I further stratified burned-unseeded and burned-seeded plots by five distance classes: <1000 m, 1000-2000 m, 2000-3000 m, 3000-5000 m, >5000 m. Burned-seeded plots distributed across three seeded areas (see Fig 2 for area C: northwest; area D: southwest; area E: east) where they were previously drill seeded with bluebunch wheatgrass Pseudoroegneria spicata) in fall 2015-2017. Bluebunch wheatgrass is a native perennial grass species widely used for post-fire restoration for its high fire tolerance (Miller et al. 2013) and relatively high establishment rate after fire (Hudson 2016).
Taking advantage of existing USGS monitoring effort, we identified 40 plots stratified by burned-unseeded (n = 15), burned-seeded (n = 15), and unburned-unseeded (n = 10) areas. Using distance from the nearest fire edge as a proxy for landscape dispersal, I further stratified burned-unseeded and burned-seeded plots by five distance classes: <1000 m, 1000-2000 m, 2000-3000 m, 3000-5000 m, >5000 m. Burned-seeded plots distributed across three seeded areas (see Fig 2 for area C: northwest; area D: southwest; area E: east) where they were previously drill seeded with bluebunch wheatgrass Pseudoroegneria spicata) in fall 2015-2017. Bluebunch wheatgrass is a native perennial grass species widely used for post-fire restoration for its high fire tolerance (Miller et al. 2013) and relatively high establishment rate after fire (Hudson 2016).
Methods. We collected plant composition data and leaf samples of bluebunch wheatgrass at all 40 plots in June 2021. Each plot was delineated by a 10 x 10 m relevé, where number of species is recorded for species richness. Occurrence of each plant taxon was measured every 0.5 m on four 25-m transects radiating from each corner of the relevé using the point-intercept method, and evenness was calculated. Leaf samples were collected from 20 individuals at each plot (n = 800), then air-dried and stored at -80F. We isolated DNA from plant tissues using a modified cetyltrimethy ammonium bromide (CTAB)-based method (Aboul-Maaty and Oraby 2019). We amplifed small quantity of DNA with polymerase chain reaction (PCR) assay using ten known primers for bluebunch wheatgrass (Bushman et al. 2008). To measure genetic diversity of bluebunch wheatgrass populations, we used a genotyping service at the Center for Genome Research and Biocomputing at the Oregon State University to obtain microsatellite data in Fall 2021. With the microsatellite data, allelic richness was calculated as a metric of genetic diversity at the population level.
Analysis. We used ANOVA to test the combined effect seeding on overall species diversity (species richness) and the genetic diversity (allelic richness) of bluebunch wheatgrass, with unburned-unseeded, burned-seeded, and burned-unseeded as explanatory variables. To test how seeding effects change with landscape dispersal, we regressed species and genetic diversity in burned-seeded/burned-unseeded plots against distance from the nearest fire edge in linear models.
Objective 3 & 4: Common Garden Experiment. Experimental design. In October 2019, we established a sagebrush steppe common garden experiment at the Northern Great Basin Experimental Range in Burns, OR. We chose a flat site with low cheatgrass (Bromus tectorum) abundance, a common invasive annual grass in Great Basin rangelands. We set up 1.5 x 3 m plots in a random block design with three rainfall treatments (n = 24 plots; 8 blocks x 3 rainfall treatments): 0% reduction for ambient rainfall, 50% reduction for moderate drought, 80% reduction for severe drought. Each plot was subdivided into twelve 25 x 25 cm subplots for nested seeding treatments (6 populations x 2 replicates). We initially nested cheatgrass competition treatments within rainfall treatments; the average densities of cheatgrass were 145.0 ± 4.0 individuals/m2 in competition plots and 7.8 ± 1.0 individuals/m2 in non-competition plots. We combined those plots as replicates of seeding treatments because there was no effect of competition on first or second year seedling survival. To reduce microsite variability from shrubs, we first manually removed aboveground biomass of shrubs to prepare the site. We constructed fixed rainout shelters that passively reduce precipitation by a constant percentage (Gherardi & Sala, 2013). Aluminum flashings were inserted 35 cm deep around the shelters to reduce surface and lateral soil flow on to the plots as well as to limit plant roots from accessing water outside the treatment. Plots were fenced with 4-ft high wire mesh that excluded cattle but not native herbivores (e.g. rabbits, gophers). In October 2019, we burned each plot with a propane blow torch to mimic post-fire conditions. To account for microclimates across rainfall treatments, we deployed soil moisture and temperature sensors (TEROS 10 and ZL6 Data Logger, Meter Group) and air temperature sensors (iButton, Maxim Integrated) in six plots. Soil moisture and temperature sensors were installed at 5 cm below the soil surface; air temperature sensors protected by radiation shields (following design by Holden et al., 2013) were installed at 10 cm above the soil surface. These sensors logged data every 2 hours year-round. In September 2020, we raked the annual plants and dug out the perennial forbs and grasses with a shovel; after removing the vegetation, we broadcasted 300 E. elymoides seeds in each subplot (one seed source per subplot). We reseeded the same density in September 2021.
Methods. In July 2021 and 2022, we measured survival rates of seedlings. Germination temperature optima of each seed source were determined by conducting germination trials in a lab at University of Oregon in November 2021. We stratified seeds in a refrigerator at a constant 4 °C for 4 weeks prior to growing them in a growth chamber. We sowed each population (25 seeds per population, 5 replicates) separately on a wet filter paper in a petri dish for 2 weeks in a constant temperature of 8, 10, 12, 15, and 20 °C. We added 2 mL of water to each petri dish at the start, and watered every 2 days to keep the filter moist. Lights were on from 6 am to 6 pm daily. Numbers of germinated seeds were counted on the 14th day. Mean proportion of germination was calculated by the dividing number of seeds germinated by the number of seeds sown. In July of 2022, we measured leaf 13C isotopic composition (δ 13C, ‰), which reflects rates of photosynthesis relative to stomatal conductance integrated over the lifespan of the leaf, as a metric of water use efficiency (dehydration avoidance; Cernusak et al., 2013). Previous work demonstrated clinal variation in carbon isotopic composition across populations of E. elymoides (Blumenthal et al., 2021). We sampled fresh leaves from two individuals of second year seedlings per subplot, and oven dried them at 60 ⁰C for 72 hours. Dried leaves were ground to powder in a ball mill, rolled in aluminum tins, and analyzed for carbon isotope composition at the Stable Isotope Lab at the University of Oregon.
Analysis. To compare the effects of seed origin and precipitation treatment (ambient, 50 % and 80 % reduction) on seedling recruitment, we built mixed models with source population and precipitation as fixed factors and block as a random factor. To compare stress tolerance strategies across seed sources, we built one-way ANOVA models with germination temperature optima and carbon isotope signature (δ 13C) of second year seedlings as dependent variables and seed sources as an explanatory variable.
Objective 1 & 2: Survey of post-fire plant communities.
I completed genotyping 800 samples (40 populations x 20 individuals) of field-collected bluebunch wheatgrass with 10 microsatellite markers.
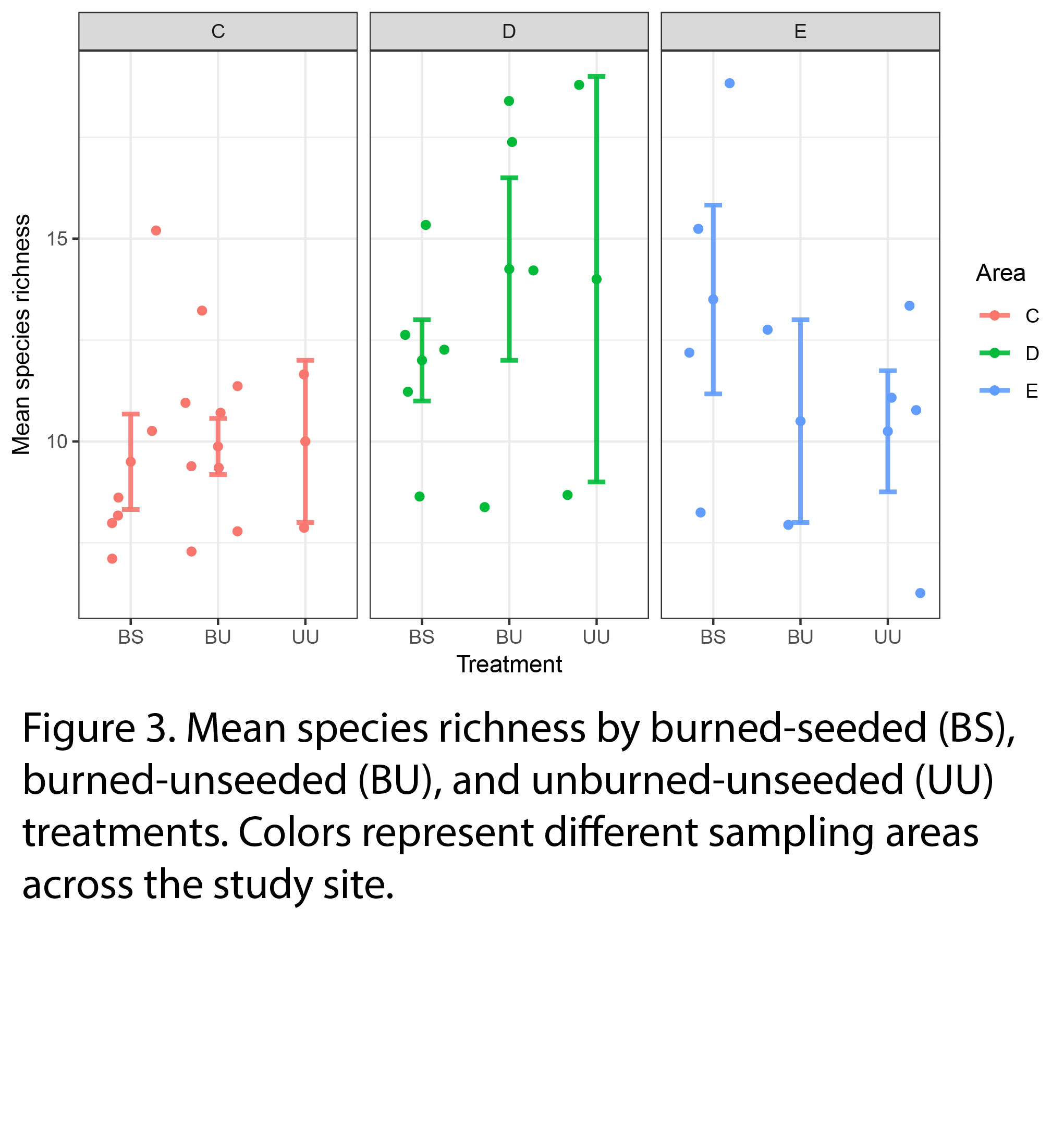 Our preliminary results show that mean species richness, a metric of species diversity, did not significantly differ by burning and seeding treatments (Fig 3), but mean species richness was significantly higher in sampling area D than area C or E.
Our preliminary results show that mean species richness, a metric of species diversity, did not significantly differ by burning and seeding treatments (Fig 3), but mean species richness was significantly higher in sampling area D than area C or E.
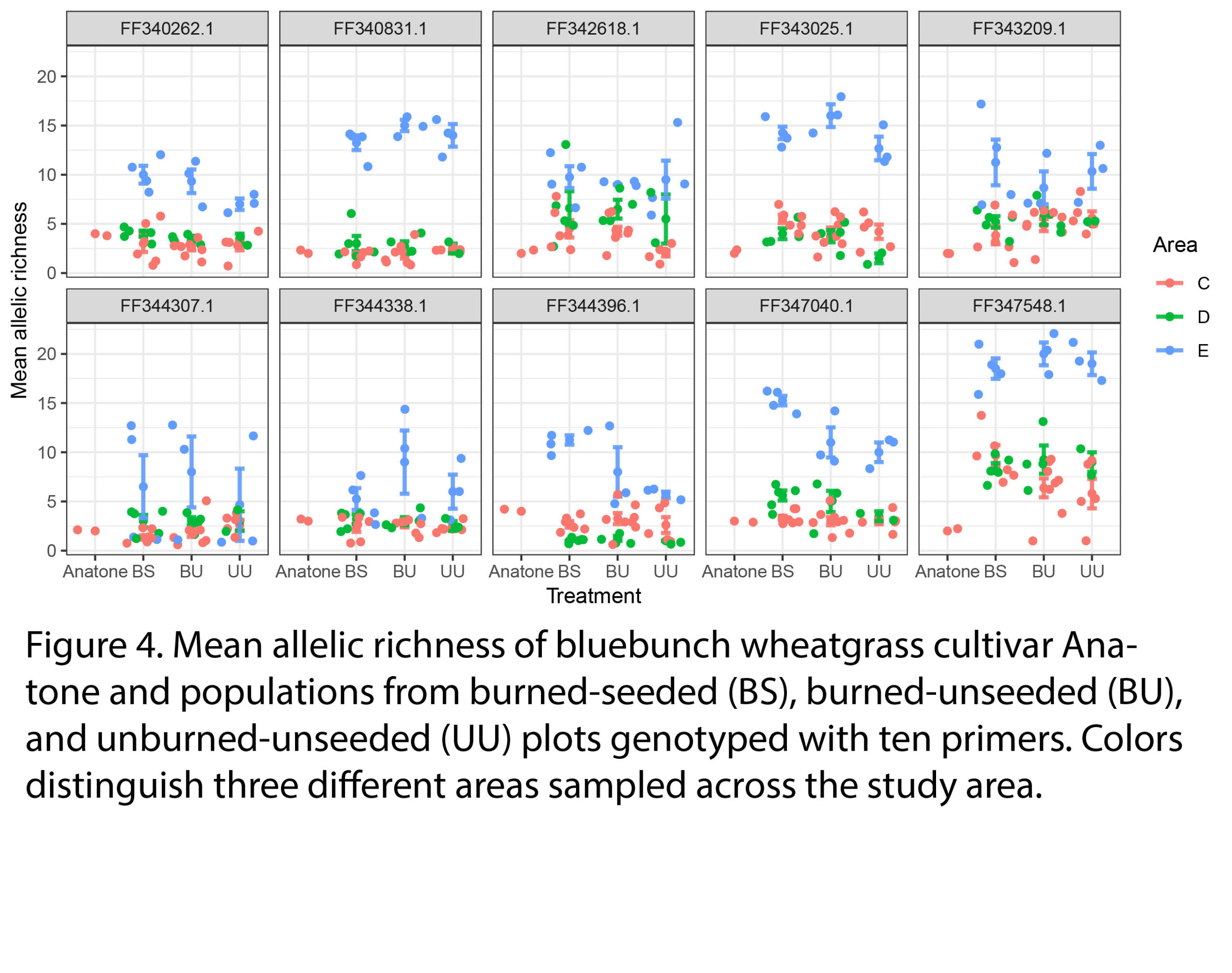 Mean allelic richness, a metric of genetic diversity, did not significantly differ by burning and seeding treatments (Fig 4), which suggests that seeded individuals are not homogenizing the genetic pool of natural populations. Mean allelic richness was significantly higher in sampling area E than area C or D.
Mean allelic richness, a metric of genetic diversity, did not significantly differ by burning and seeding treatments (Fig 4), which suggests that seeded individuals are not homogenizing the genetic pool of natural populations. Mean allelic richness was significantly higher in sampling area E than area C or D.
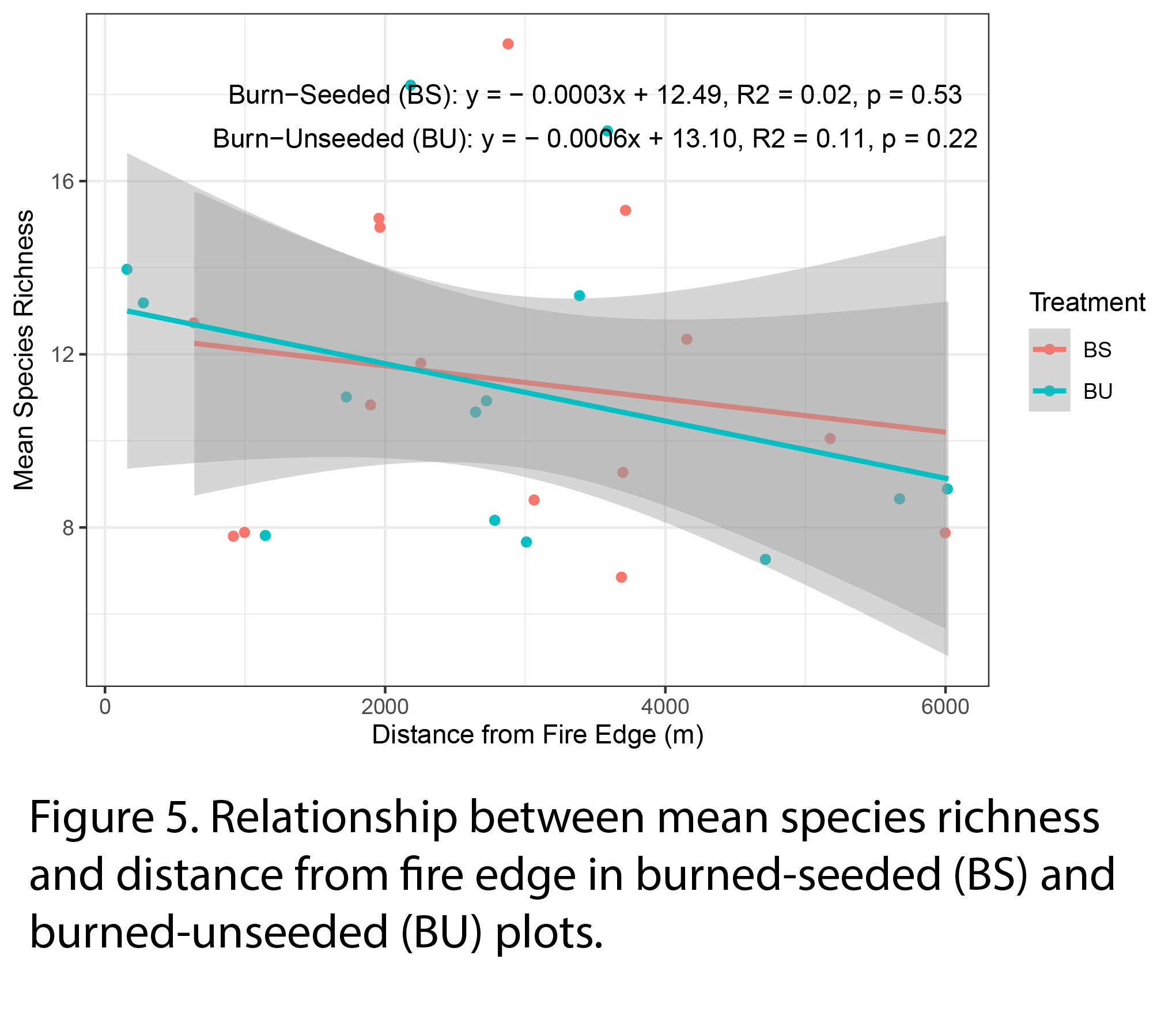 Mean species richness slightly decreased with distance from fire edge in both seeded and unseeded burned areas but their relationships were non-significant (Fig 5).
Mean species richness slightly decreased with distance from fire edge in both seeded and unseeded burned areas but their relationships were non-significant (Fig 5).
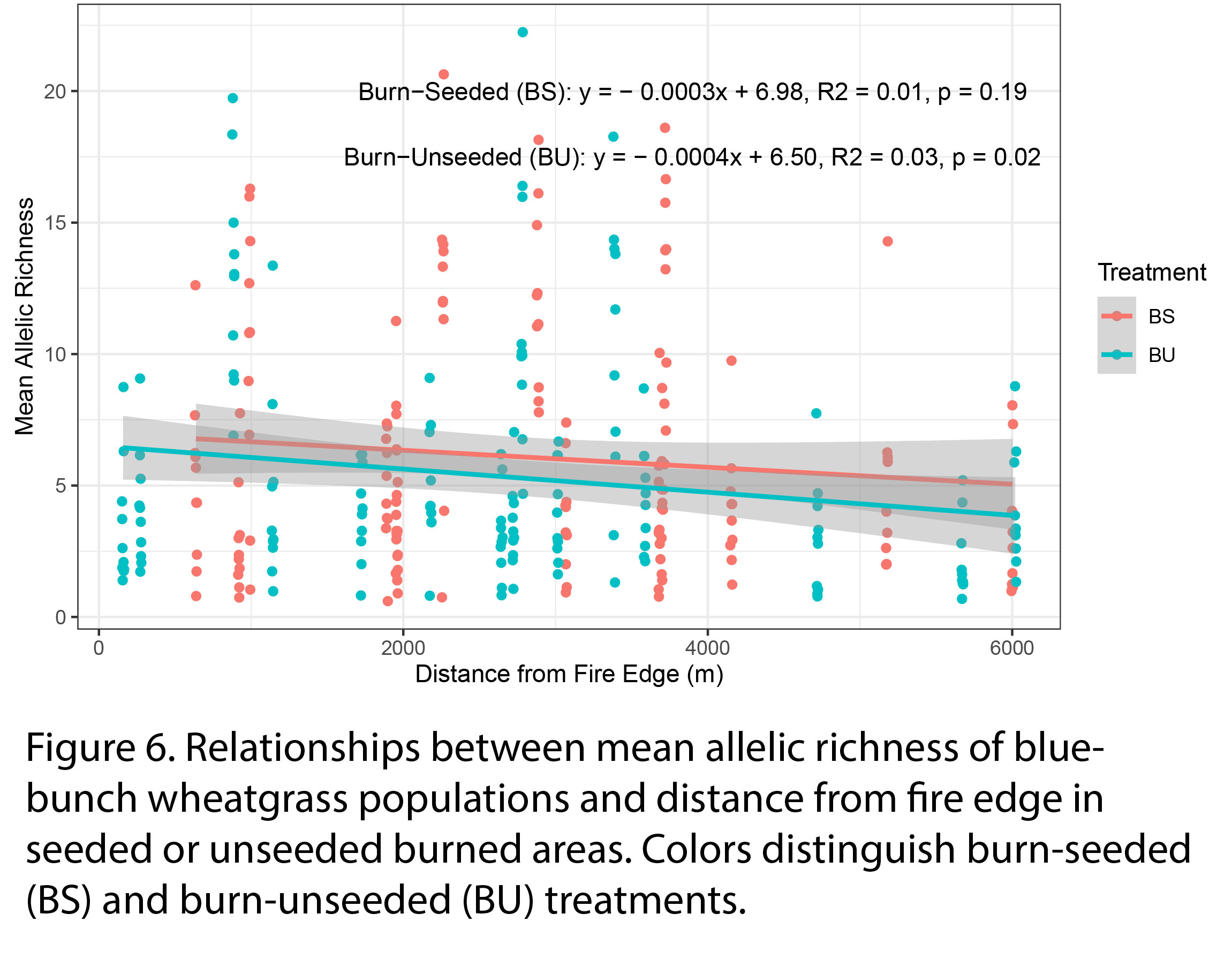 While mean allelic richness slightly decreased with distance from fire edge in burned-unseeded area (p = 0.02), we observed no strong relationship between mean allelic richness and distance from fire edge in burned-seeded areas (Fig 6). Our results may suggest that natural recovery from seedbank or pre-existing populations that survived the burn are maintaining the genetic diversity of bluebunch wheatgrass populations, and seed dispersal from neighboring populations may be longer ranged than previously thought. Future research should quantify the seed dispersal range (kernel) of bluebunch wheatgrass to understand how natural recovery is occurring on this landscape.
While mean allelic richness slightly decreased with distance from fire edge in burned-unseeded area (p = 0.02), we observed no strong relationship between mean allelic richness and distance from fire edge in burned-seeded areas (Fig 6). Our results may suggest that natural recovery from seedbank or pre-existing populations that survived the burn are maintaining the genetic diversity of bluebunch wheatgrass populations, and seed dispersal from neighboring populations may be longer ranged than previously thought. Future research should quantify the seed dispersal range (kernel) of bluebunch wheatgrass to understand how natural recovery is occurring on this landscape.
Objective 3 & 4: Common Garden Experiment.
Seeding and Rainfall Treatments
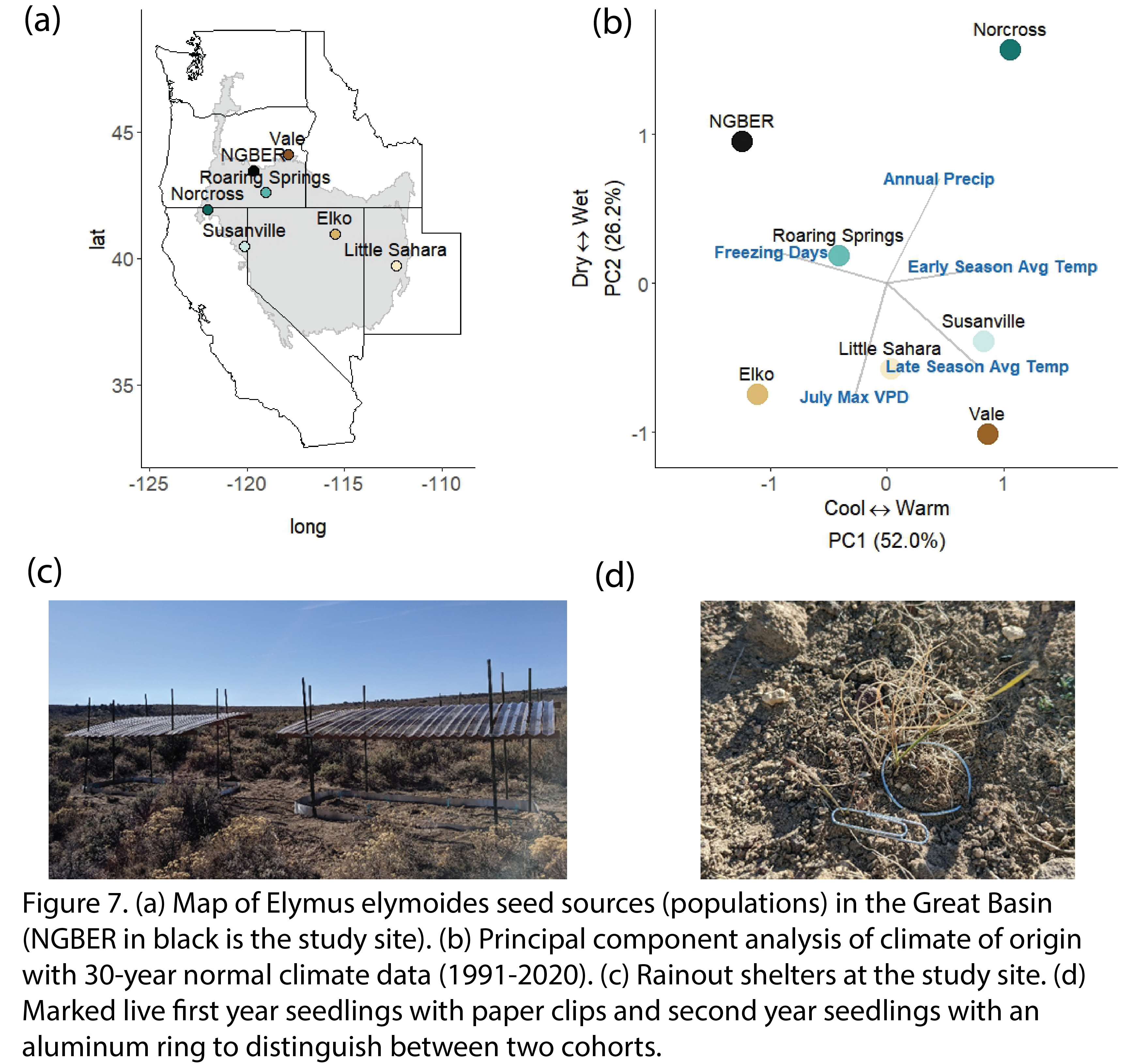
Bottlebrush squirreltail (Elymus elymoides) seeds were sourced from six sites (Fig 7a) along an aridity gradient across the Great Basin (Fig 7b). We conducted a common garden experiment in a sagebrush steppe in Oregon to evaluate the seedling recruitment responses to climate-adjusted provenancing and rainfall treatments that varied in soil moisture (Fig. 7c). We distinguished first year seedlings from second year seedlings by using visible phenology cues (i.e., new recruits had seed glumes still attached or no standing dead biomass) and aluminum rings with different widths (Fig. 7d).
Treatment Effects on Microclimate
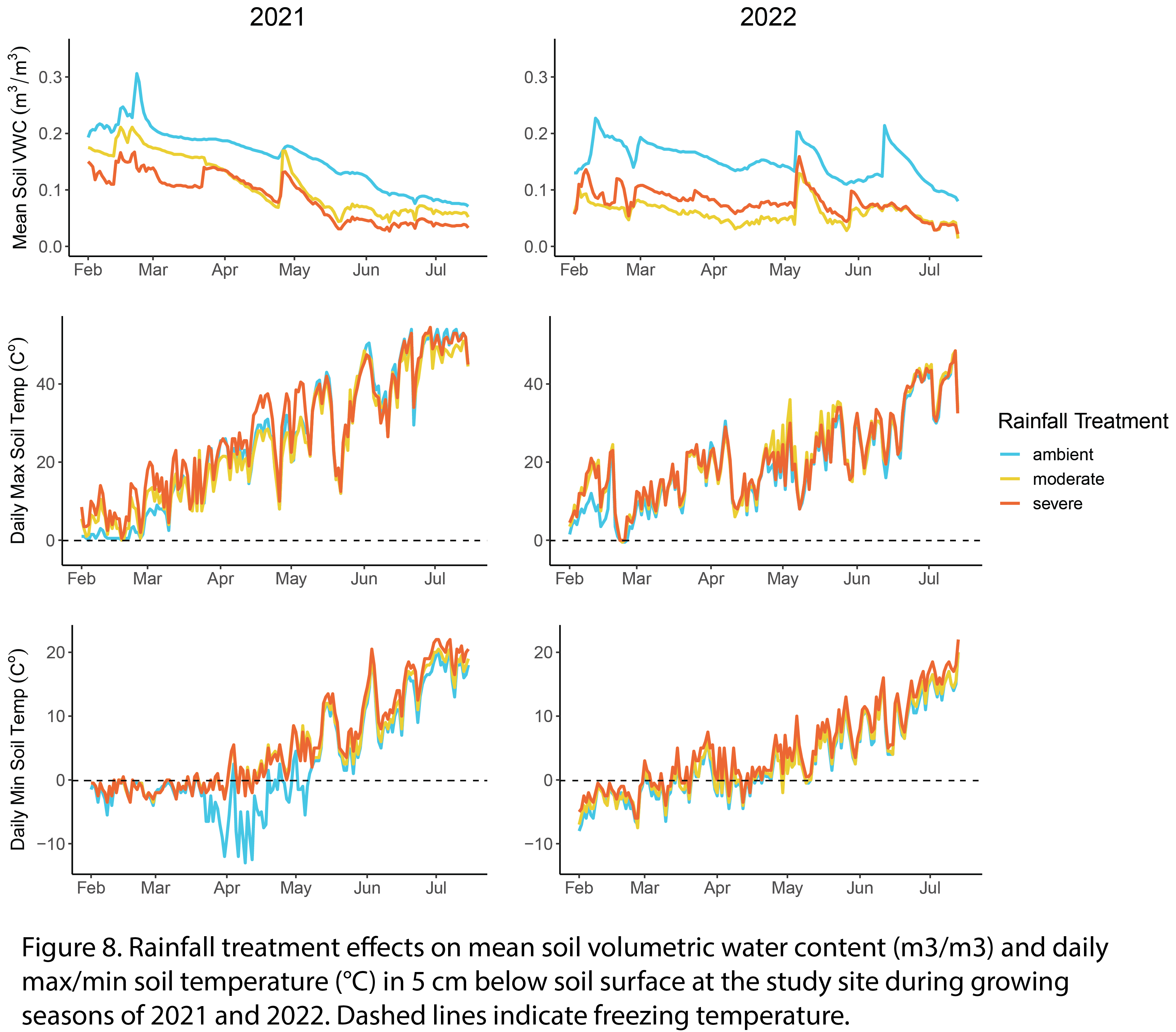
Rainout shelters effectively reduced soil moisture in top 5 cm of the soil surface by 36% and 62% in moderate and severe drought plots, respectively (Fig. 8). Soil temperatures in top 5 cm of the soil surface were on average 0.7 °C and 1.5 °C warmer in moderate and severe drought plots compared to ambient rainfall plots (Fig. 8). The 2021 growing season began with a wet and particularly cold spring (below zero daily min soil temp in February to May) followed by a heat dome in the summer (above 40 ⁰C daily max soil temp in June and July; Fig. 8). The 2022 growing season was overall wetter, with two rainfall events in May and June (Fig. 8).
Seedling Performance
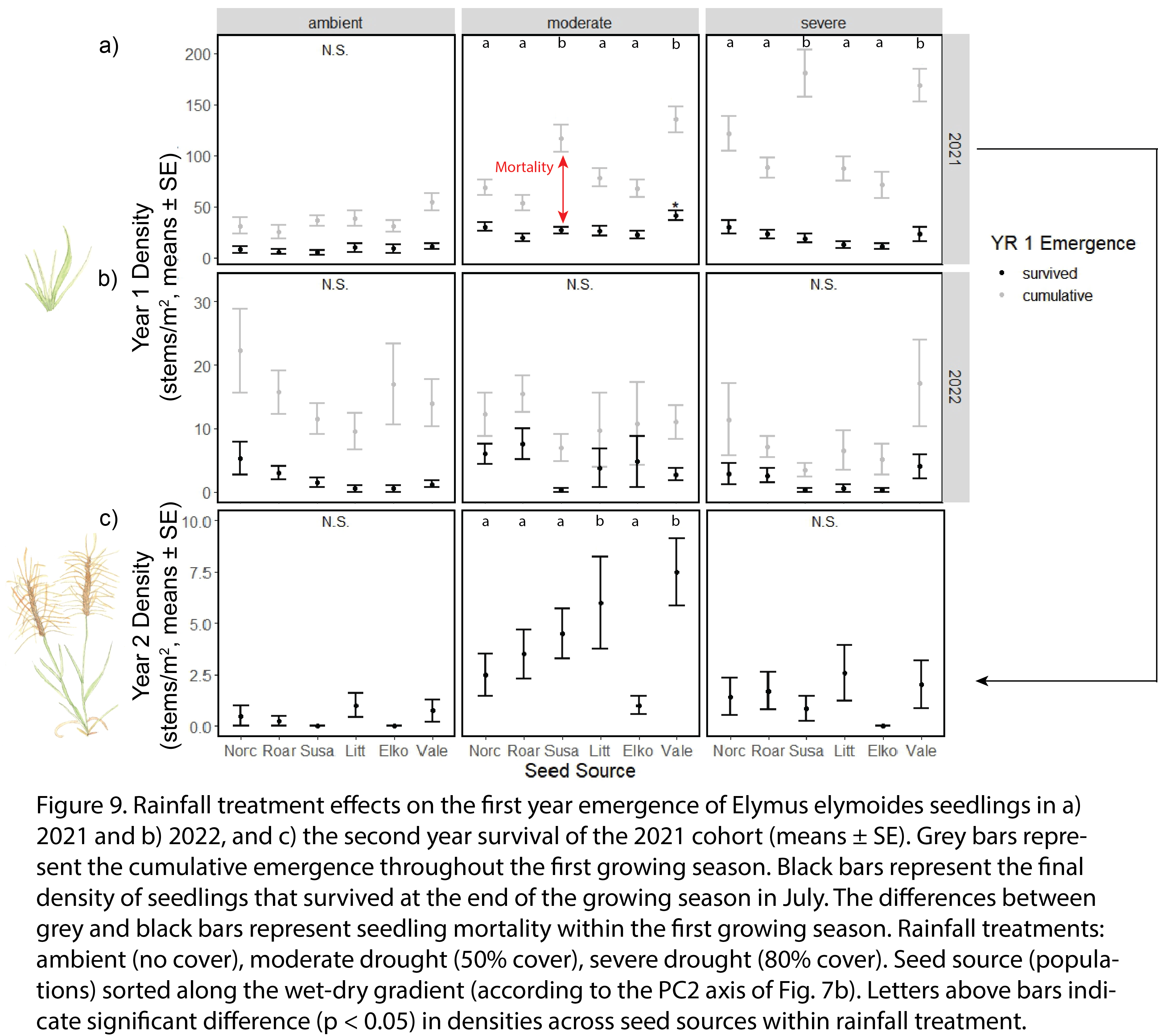
The cumulative emergence were 80.8 ± 3.8 and 11.6 ± 1.0 seedlings/m2 in 2021 and 2022, respectively, out of a seeding rate of 4,800 seeds/m2. Of the 2021 cohort, Vale and Susanville had significantly higher cumulative emergence than other seed sources in moderate and severe drought conditions (Fig. 9a). We observed higher cumulative emergence in drought plots than ambient plots in 2021 (Fig. 9a), but lower cumulative emergence in drought plots than ambient plots in 2022 (Fig. 9b).
The final densities of seedlings at the end of the first growing season were 12.2 ± 0.3 and 1.5 ± 0.1 seedlings/m2 in 2021 and 2022, respectively. Of the 2021 cohort, seeds from Vale had significantly higher densities of first year seedlings than other seed sources in moderate drought conditions (Fig. 9a). First year seedling survival rates in 2021 and 2022 were half in severe drought (0.17 ± 0.02; 0.17 ± 0.04) compared to moderate drought (0.33 ± 0.02; 0.34 ± 0.07).
The final densities of seedlings at the end of the second growing season - a benchmark of restoration success - in ambient rainfall, moderate drought, and severe drought conditions were 0.4 ± 0.2, 4.2 ± 0.6, and 1.4 ± 0.4 seedlings/m2, respectively. Two populations from warmer and drier climate zone – Vale and Little Sahara – had higher second year seedling survival in moderate drought conditions than the more ‘local’ seed source (Roaring Springs) in terms of both geographic distance and environment (Fig. 9c); this result supported our hypothesis that climate-adjusted provenancing increases seedling performance.
Stress Tolerance Strategies
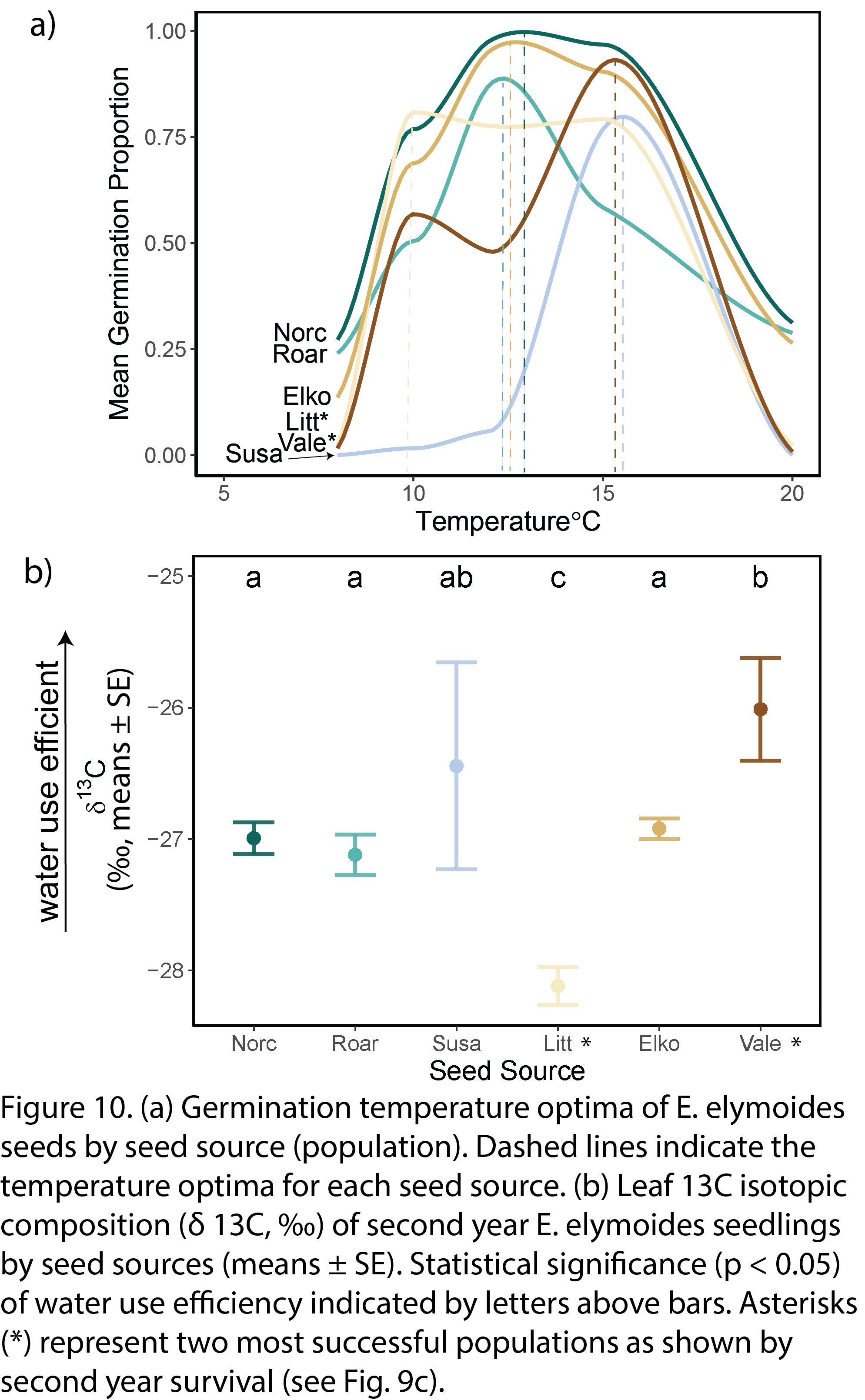
Germination temperature optima varied by seed sources (Fig. 10a): Little Sahara at 10 °C, Norcross, Elko, and Roaring Springs at 12 °C, and Vale and Susanville at 15 °C. Water use efficiencies measured by stable carbon isotopic compositions (δ 13C) also varied by populations (Fig. 10b). In particular, Vale was most water use efficient, while Little Sahara was least water use efficient. Vale seeds with delayed germination and high water use efficiency avoided frost mortality and tolerated drought, while Little Sahara seeds with early germination and low water use efficiency capitalized on two water events that occurred in the second growing season. Understanding these differences in stress tolerance strategies among populations within species is critical for creating seed mixes that increases the adaptive capacity of the restored population.
References
Aboul-Maaty, N. A. and H. A. Orgaby. 2019. Extraction of high-quality genomic DNA from different plant orders applying a modified CTAB-based method. Bulletin of the National Research Centre 43:25.
Baughman, O. W., A. C. Agneray, M. L. Forister, F. F. Kilkenny, E. K. Espeland, R. Fiegener, M. E. Horning, R. C. Johnson, T. N. Kaye, J. Ott, J. B. St. Clair, and E. A. Leger. 2019. Strong patterns of intraspecific variation and local adaptation in Great Basin plant revealed through a review of 75 years of experiments. Ecology and Evolution 9:6259-6275.
Craine, J. M., T. W. Ocheltree, J. B. Nippert, E. G. Towne, A. M. Skibbe, S. W. Kembel, and J. E. Fargione. 2012. Global diversity of drought tolerance and grassland climate-change resilience. Nature Climate Change 3: 63-67.
Damschen, E. I., D. V. Barker, G. Bohrer, R. Nathan, J. L. Orrock, J. R. Turner, L. A. Brudvig, N. M. Haddad, D. J. Levey, and J. J. Tewksbury. 2014. How fragmentation and corridors affect wind dynamics and seed dispersal in open habitats. PNAS 111(9): 3484-3489.
Farquhar, G. D., J. R. Ehleringer, and K. T. Hubick. 1989. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40: 503-537.
Foster, B. L., and D. Tilman. 2003. Seed limitation and the regulation of community structure in oak savanna grassland. Journal of Ecology 91(6): 999-1007.
Hoffmann, A. A., and M. J. Hercus. 2000. Environmental stress as an evolutionary force. BioScience 50(3): 217-226.
Hudson, T. D. 2016. Seeding after fire. Washington State University Extension. https://s3.wp.wsu.edu/uploads/sites/2063/2016/03/Seeding-After-A-Fire.pdf.
Ives, A. R., and S. R. Carpenter. 2007. Stability and diversity of ecosystems. Science 317: 58-62.
Leger, E. A. and O. W. Baughman. 2015. What seeds to plant in the Great Basin? Comparing traits prioritized in native plant cultivars and releases with those that promote survival in the field. Natural Areas Journal 35: 54-68.
Leger, E. A., D. Z. Atwater, and J. J. James. 2019. Seed and seedling traits have strong impacts on establishment of a perennial bunchgrass in invaded semi-arid systems. Journal of Applied Ecology 56(6): 1343-1354.
Miller, R. F., J. C. Chambers, D. A. Pyke, F. B. Pierson, and C. J. Williams. 2013. A review of effects of vegetation and soils in the Great Basin region: response and ecological site characteristics. USDA Forest Service RMRS-GTR-308.
Nathan, R. and H. C. Muller-Landau. 2000. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in Ecology & Evolution 15(7): 278-285.
Research outcomes
Forecasted climate change trends toward increased ecological drought in rangeland ecosystems will likely have a large effect on future restoration efforts. We provide a proof of concept for the climate-adjusted provenancing approach to seed sourcing as a bet-hedging strategy to improve restoration outcomes in warmer and drier environments in the future. Our combined approach of a manipulative field experiment and a targeted trait screening enabled us to understand how seedlings with diverging stress tolerance strategies perform under variable environmental conditions. Sparse data are available to describe the exact environmental conditions likely to cause mortality for restoration target species or sources within species. Further research on intraspecific variation in frost tolerance of seedlings is needed to select seeds precisely suitable for the environmental condition of the restoration site. Our results suggest that no one seed source is a silver bullet for seeding success, and there is merit to mixing local seeds with seeds pre-adapted to future conditions.
Education and Outreach
Participation summary:
To reach natural resource managers in the Bureau of Land Management district offices across the Northern Great Basin, I hosted two zoom meetings to share and discuss my findings with them on November 8th and 9th, 2022. The format of the meeting was one hour of presentation and one hour of discussion on challenges and opportunities for modifying seed sourcing practices and managing invasive annual grasses. I also shared research updates at the virtual seminar event hosted by the Eastern Oregon Agricultural Research Center in Burns, Oregon on September 29, 2021, and at the Society of Range Management Pacific Northwest Section's "Home on the Range" webinar on October 5, 2021. Recording of the talk is archived here: https://www.youtube.com/watch?v=HZl1-VfaTYw&list=PLl12IcBpmhGsWENxTxuhqATJAheGDYHeP&index=4
I mentored three undergraduate researchers at University of Oregon. I helped them secure research funding and guided them through
their independent research projects.
As a leader of the student chapter of the Society for Ecological Restoration at University of Oregon, I organized monthly volunteer events at local parks and on campus in the 2021-23 academic year for graduate and undergraduate students to gain hands-on experience on habitat restoration. I also facilitated a four-part virtual webinar on prescribed fire and post-fire restoration in the Pacific Northwest through the student chapter of the Society for Ecological Restoration at University of Oregon on February 15-16 & 22-23, 2022. Recordings of the webinar is archived here: https://blogs.uoregon.edu/uoser/2022/03/
I gave two public presentations: an annual conference for the Society for Ecological Restoration Great Basin Chapter on April 27th, 2023 and an annual conference for the Ecological Society of America on August 9, 2023.
I published two articles (see below), one in review, and one more in progress.
Aoyama, L., E. A. Shaw, C. T. While, K. N. Suding, and L. M. Hallett. 2023. Functional diversity buffers biomass production across variable rainfall conditions through different processes above- vs belowground. Functional Ecology 00: 1-15. https://doi.org/10.1111/1365-2435.14394
Aoyama, L., E. Cook, and L. M. Hallett. 2022. Intraspecific variation in native grass seedling response to water stress depends on the annual grass invasion context. Restoration Ecology e13816. https://doi.org/10.1111/rec.13816
I created a website where I publicly shared my research progress: https://www.linaaoyama.com/
Total of 23 natural resource managers in the Bureau of Land Management district offices in Twin Falls, Burns, Vale, and Boise attended the zoom meetings. They learned that seed sourcing for post-fire restoration is an important decision they make in reshaping the plant communities for rangeland resilience. While the current reality is using any seeds that are available, the managers were keen on modifying their seed sourcing practices if there is reliable access to seeds from variable populations of the target species.
From my mentorship, three undergraduate students gained fieldwork and lab experiences, developed confidence to conduct independent research, and completed their honors theses.
Native plant restoration projects were attended by total of attended by total of 18 undergraduate students and 5 graduate students. Eight speakers participated in the the speaker series including an elder from the Karuk Tribe, fire ecologists from USFS, Oregon State University, and non-profits. Total viewers were 64 people.
Total participants for the SER talk: 51 people. Total participants for the ESA talk: 40 people.
Functional Ecology article's full text views: 795. Restoration Ecology article's full text views: 244.