Final Report for ONE13-187
Project Information
Building and maintaining soil organic matter is central to the productivity of vegetable farms. One of the challenges for stockless vegetable farms is the need to offset losses of soil organic matter and nutrients attendant with tillage and the selling of produce. To date, many vegetable growers import compost from off-farm sources as their reliable source of additional soil organic matter.
In this study we examined the potential use of alder chips as a soil conditioner in intensive vegetable production systems over a three-year rotation of tomato, Brussels sprouts, and black beans. We compared yields, weed biomass, and changes soil quality parameters based on chips either incorporated into the soil or as mulch, in addition to a treatment with incorporated chips made from spring harvested, leafy branches. Control treatments were compost and no compost. For each year, the treatment plots received a new application of chips.
Surprisingly, there were no significant differences among alder treatments on final yields for tomatoes, Brussels sprouts and beans. Furthermore, only the tomatoes showed a significantly higher yield with compost and this difference only became significant after a sustained heat wave in early September that slowed growth and appeared to be exacerbated by low tissue potassium content of all treatments except for the compost control.
Soil and weed responses, in contrast, changed over the three years of the experiment. The Chip + mulch treatment received twice the amount of carbon input and there was a noticeable increase in the sifted-fraction of soil organic matter (1.4%) in contrast to the treatment with no soil conditioner with an overall decrease of 3% over the three years of the experiment. The incorporated chip, mulch, and spring chip treatments lost approximately 1% SOM in the sifted-fraction over the same time period. Degree of compaction as measured with a penetrometer, paralleled these patterns of SOM with the highest compaction in the no soil conditioner treatment and lowest across all chip treatments.
Not surprisingly, weed biomass varied with both crop and treatment. Overall weed biomass was highest with Brussels sprouts while the beans had consistently lowest. Overall, the mulched treatments had an order of magnitude fewer weeds. The spring chip appeared to have an allelopathic effect, with significantly lower weed biomass in Brussels sprouts and beans. This allelopathic effect also of spring chips was suggested by the slow and poorer germination of the direct seeded beans.
Overall, winter chipped alder appears to be a viable source for building SOM, providing a balanced macro- and micro-nutrient supply, and for effective weed suppression. Direct seeding into the coarse chip mulch of preceding years can be challenging but did not have a significant yield effect in the beans. The spring mulch behaved differently in soils with higher microbial activity, different abundance of bacteria and fungi in the soil food web, and allelochemical suppression of annual weeds.
Regrowth of cut alder varied depending on sun exposure. In full sun, basal branches for the cut stump reached sexual maturity within 3 years. A three-year to five-year coppice cycle can provide branches of appropriate size for ramial chip production in addition to providing ecological services such as a carbon and nitrogen fixation in a buffer strip as well as important habitat for a broad range of organisms from worms, cottontail, and woodcock.
Alder is often the focus of various roadside eradication schemes that include burning and chipping. Winter chipped alder can be a viable off or on-farm source of soil organic matter and may particularly well suited for no-till vegetable production systems that focus on the use of large volumes of organic matter. These winter chips also a good substrate for edible mushroom growth such as the wine cap, another potential source of income in chip-based system.
Introduction:
Coppicing of hardwood species has a long tradition in the UK (Harmer 1995) and once again is being developed throughout Europe for biofuels (Jørgensen et al. 2005), wildlife management (Fuller and Warren 1993), and wood from crafts such as thatching and basket making (Harmer 1995). Coppicing of shrubs found on farm pasture and woodlots might be one innovative solution for New England farmers to establish a perpetual regimen of carbon and nutrient harvesting that can be used to fuel soil formation and to transfer mineral nutrients from surrounding woody edges into agricultural production areas. Historically, this kind of nutrient transfer was mediated through the nighttime collection of manure from animals that browse in forest during the day. This proposal is a stockless form where coppicing and chipping are mechanical replacement for mouths with grinding teeth, and has the potential advantage of transferring more organic carbon and nutrients directly to the soil faunal and microbial decomposer communities.
In this project we examine whether Maine farmers might add energy and plant nutrients, and build soil organic matter for vegetable production through the use of currently underutilized on-farm resources of ramial wood (small branches of hardwood species of < 3 inches in diameter) from forests, woodlots, and pasture edges. This study focused on the chipping of the nitrogen-fixing alder for ramial wood chips (RWC). In Maine, alder (Alnus spp) is often perceived as an invasive species of pasture or a trash tree in forestry. The ability of alder to regenerate from cutting, however, has been utilized in habitat management for the woodcock (Williamson 2010). The twenty-year rotations for bird habitat management in Maine suggest that the local species are amenable to coppicing. European studies conducted on Alnus incana (one of our local species) suggest that coppicing on a 3 to 5 year cycle would produce a resilient and healthy stand that could be used by farmers on a long term basis (Uri et al 2011).
Previous studies on traditional farming systems in China (GIAHS 2012) , Nepal(Pulamte 2009), India (Rathore et al. 2010) and Mexico (CEDICAM 2012) have demonstrated the importance of alder biomass for the building and maintenance of long term fertility of agricultural systems (Tarrant and Trappe 1971, Brown 2012). Tissue analyses indicate that leafy twigs and small branches contain approximately 1.5 % nitrogen and have a C/N ratio of 25:1 (Uri et al 2003, Tarrant and Trappe 1971). Studies also have demonstrated that alder easily decomposes and, due to lignin forms in young leaves and branches, tends to form relatively stable humus (Seo et al. 2011 , Sharma 1993).
In Maine, use of alder ramial wood chips has been documented only in fruit tree orchards (Currier 2007) and not used for vegetable production despite impressive findings in a fifteen-year study in New York (Free 1971), in Canada (Lemieux 1996), and more recently in Senegal (2002). In this project, we focus on alder because of its abundance and often weedy nature on farms, its favorable C:N ratio (which may solve the typical tie-up problem seen with other woody tissues), its rapid decomposition, and its potential as a local, renewable source of energy for the soil decomposer communities and of soil carbon and nitrogen. Coppicing and chipping also are an easy way to move other minerals from the forest and forest edge into the more nutrient-demanding cropping areas. All of these contribute the sustainability of the farm by decreasing the dependence on off-farm inputs and instead using local, almost entirely renewable resources.
Breland TA Eltun R (1999) Soil microbial biomass and mineralization of carbon and nitrogen in ecological, integrated and conventional forage and arable cropping systems. Biology and Fertility of Soils 30: 193-201
Brinton WF, Sparda AR Haney RL (2011) Solvita: a new, rapid soil test for active soil carbon and N-mineralization. Woods End Laboratories
Brown K (2012) Beyond nitrogen: how is red alder growth affected by mineral nutrient supply. BC Forest Professional 1: 18-19
Caron C, Lemieux G, Lachance L (1998) Regenerating soils with ramial chipped wood. Publ no 83, Laval University
Caron C (2006) Connecting with the terrestrial ecosphere. The Maine Organic Farmer and Gardener CEDICAM http://www.ecotippingpoints.org/our-stories/indepth/mexico-oaxaca-community-reforestation-mixteca-region.html accessed 9.01.2012
Chapin FS Kedrowski R (1983 ) Seasonal changes in nitrogen and phosphorous fractions and autumn retranslocation I evergreen and deciduous taiga trees. Ecology 64: 376-391
Currier A (Fall-winter 2007-2008) Using Ramial chipped wood to improve fertility in a fruit tree nursery The Maine Organic Farmer and Gardener
DeBell DS Radwan MA (1979) Growth and nitrogen relations of coppiced black cottonwood and red alder in pure and mixed plantings. Botanical Gazette 140: S97-S101
Free GR (1971) Soil management for vegetable production on Honeoye soil with special reference to the use of hardwood chips. New York’s Food and Life Sciences Bulletin 2: 1-20
Fuller RJ Warren MS (1993) Coppiced woodlands: their management for wildlife. 2nd edition. JNCC, Peterborough GIAHS --Globally Important Agricultural Heritage Systems http://www.giahs.org/sites/south-east-asia/alder-based-rotation-and-intercropping-yunnan-china/detailed-information/en/ accessed 9.01.2012
Harmer R (1995) Management of coppice stools. Research Information Note 259, The Forestry Authority, UK
Hurd TM Raynal DJ Schwintzer (2001) Symbiotic N2 fixation of Alnus incana ssp. rugosa in shrub wetlands of the Adirondack Mountains, New York, USA. Oecologia 126: 94-103
Jørgensen U Dalgaard T Kristensen ES (2005) Biomass energy in organic farming—the potential role of short rotation coppice. Biomass and Bioenergy 28: 237- 248
Lemieux G (1996) The hidden world that feeds us: the living soil. Publ 59b Laval University, Quebec Nordh NE
Nordh NE Verwijst T (2004) Above-ground biomass assessments and first cutting cycle production in willow (Salix sp.) coppice—a comparison between destructive and non-destructive methods. Biomass and Bioenergy 27: 1-8 Peichl M
Leava NA Kiely G (2012) Above- and belowground ecosystem biomass, carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grassland. Plant Soil 350: 281-296
Pulamte L (2009) Linkage between indigenous agriculture and sustainable development—evidences from two hill communities in northeast India. Proceedings GLOBELICS, 7th International Conference, 6-8 October, Dakar, Senegal.
Seo KW Heo SJ Son Y Noh NJ Lee SK Yoon CG (2011) Soil moisture condition and soil nitrogen dynamics in a pure Alnus japonica forest in Korea. Landscape Ecol Eng 7:93-99
Sharma E (1993) Nutrient dynamics in Himalayan alder plantations. Annals of Botany 72: 329-336
Soumare MD, Mnkeni PNS Khouma M (2002) Effects of Casuarina equisetifolia composted litter and ramial-wood chips on tomato growth and soil properties in Niayes, Senegal. Biological Agriculture and Horticulture 20: 111-123
Tarrant RF Trappe JM (1971) The role of Alnus in improving the forest environment. Plant Soil, special volume, pp 335-348
Uri V, Tuulus H Lohmus K (2003) Nutrient allocation, accumulation and above-ground biomass in grey alder and hybrid alder plantations. Silva Fennica 37(3): 301-311
Uri V Lohmus K Mander U Ostonen I Aosaar J Maddison M Hilmisaari HS Augustin J (2011) Long-term effects on the nitrogen budget of a short-rotation grey alder (Alnus incana(L.) Moench) forest on abandoned agricultural land. Ecological Engineering
Verwijt T Telnius B (1999) Biomass estimation procedures in short rotation forestry. Forest Ecology and Management 121: 137-146
Wang L Gruber S Claupein (2011) Effects of woodchip mulch and barley intercropping on weeds in lentil crops. Weed Research 52: 161-168
Williamson S (2010) American Woodcock: Habitat Best Management Practices for the Northeast. U.S.DA, Natural Resources Conservation Service. Washington, DC
Objective 1: Assess changes in crop yields and soil organic matter that occur over a three year rotation as the result of the application of alder woodchips as an amendment to current grower practices and as a possible substitute for off-farm compost.
Objective 2: Provide farmers guidelines for coppicing alders and other hardwoods on the edge of their farm properties
Cooperators
Research
YEAR 1 (2013)
Alder was harvested from fence lines on Peggy Rockefeller Farm (PRF) early in the winter and stockpiled for processing at the time of transplanting (Figure 1). Over 2 yards of coarse ramial wood chips (RWC) and less than one yard of coarse spring alder were chipped all in one day with a rented chipper (Figure 2). Brush piles were chipped at the time of tomato crop establishment and analyzed along with initial field soil samples. Eleven of the cut individuals were marked for re-growth studies (FIGURE 1).

Figure 1 Aerial view of alder transect and growth after harvest. 
Figure 2 Coarse mix of alder ramial wood chips. Inset: piles of winter and spring chipped alder
.
Field experiments were conducted on the two farms, Peggy Rockefeller Farm of College of the Atlantic (Tunbridge-Lyman, Hancock County, average GDD 3740) and Daybreak Farm (Boothbay silt, Knox County, average GDD - 4566). Three randomized, replicate beds (10 ft by 2.5 ft) were established for each of the following treatment (FIGURE 3).

Figure 3 Tomato trials on DBK and PRF
Control: compost
Treatment 1: no amendments (“none”)
Treatment 2: alder chip incorporated (“chip”)
Treatment 3: alder chip and alder mulch (“chip+mulch”)
Treatment 4: alder mulch (“mulch”)
Treatment 5: spring alder chip with leaves (“spring chip”)
The control for all treatments was based on tomato production practices used on COA’s vegetable farm (Beech Hill Farm), which included pro-gro and crab meal at time of planting. Treatment 1 had no additional organic matter (no OM) and allows for a direct comparison of with and without compost while treatment three allows for a comparison of with and without alder. Comparisons between 3 and 4 allows for test of the role of mulch with and without incorporated chips. While treatment five allows for a direct comparison of chips harvested in spring vs winter.
On Daybreak Farm, three replicates of treatments 1 and 2 in addition to the compost control were followed through the growing season. All five treatments were followed at Peggy Rockefeller Farm and were designed to look at the effects of alder chips as a substitute for compost and as mulch. For each replicate strip with an alder treatment, twenty pounds of chips were inoculated with half oz. forest duff in order to increase fungal abundance (Caron 1998), and then incorporated into the top 2 to 3 inches. Mulch treatments received an additional 20 lbs (=1 bushel) of inoculated chips. These application rates are following methods used by Free (1971). Each control plot (with and without compost) also received the forest duff inoculant.
All tomato plants were started in the College of the Atlantic greenhouse in Fort V growing mix on 8 May and transplanted into 4- inch peat pots 14 May. Daybreak (DBK) and Peggy Rockefeller (PRF) farms received transplants on 8 June and transplanted into the field on 21 June (Figure 4). At the time of transplanting, Daybreak basal diameters were significantly larger (p=0.07 for Bellstar and p=0.002 for Orange blossom). By the end of July, there was no difference between the two farms for orange blossom and belle star. Tomatoes were trellised and grown at 1.5-ft spacing, with approximately 7 plants per ten ft strip. To avoid edge effects, 3 randomly selected plants from center of each replicate were marked and followed throughout the growing season.
Three growth estimates (basal diameter and height) were made between time of transplanting and the first harvest in August (Figures 5 and 6). Fruit number and weight were followed from mid-August until the end of the growing season (early Sept for DBK and early October for PRF). Leaf tissue analysis was taken in late August of three plants per rep per treatment. The last harvest for DBK was at the end of August with the onset of late blight in all treatments while the last harvest for PRF was the fifth of October. During July, August, and September, fruit from the 3 designated individuals within treatment blocks will be collected and weighed two to three times a week on both farms.
In addition to yields, the variation in total biomass of in-row weeds were harvested and weighed in July and August.

Figure 4. Mulch treatment

Figure 5. Field measurements of crop and alder coppice

Figure 6. Treatments in center aisle ( mulch, spring, compost) in mid-June and early July.
YEAR 2 (2014)
In the second year, alder chip treatments were repeated on the Peggy Rockefeller Farm experimental plots. . Prior to the spring growing season, soil samples for all 18 trial plots were pulled for basic soil tests. Experimental plots locations remained same and the tomatoes were followed up with a Brussels sprouts rotation. Alder branches less than 3” in diameter harvested in the winter and fresh, spring alder with leaves were coarsely chip one week prior to the transplanting of Brussels sprouts in mid-June.
All transplants (var. Diablo) were started in the College of the Atlantic greenhouse in blocks of Fort V growing mix in late May and were potted up into 4-inch pots. At the time of transplanting (28 June), all plants received pro-gro and crab meal, and drip irrigation was installed. Based on the spring soil analyses, all treatments except for the compost control received sul-po-mag in order to compensate for the decline in potassium availability over the 2013 growing season.
Cabbage white butterfly was a common visitor on the Brussels sprouts after the removal of the floating row cover at the end of July. In September, 17 students worked together to sample the top 5 inches of each stalk for the number of aphids (all 0), the number of cabbage white caterpillars, the total number of leaves in the top 5 inches and the number of leaves with any caterpillar damage. A damage code was assigned to each plant to represent the severity of caterpillar damage: 0=no damage, 1= 25% of the leaf surface removed, 2=-50%, 3=75% of the leaf removed. This code was a visually determined average of the leaves in the sample area (leaves coming from the top 5" of stalk).
Plant basal diameter was recorded twice during the growing season. Inter-row weed biomass and diversity were sampled mid-July and August. Final harvest occurred in mid-November prior to significant freezing temperatures. Total above ground biomass and total fresh weight of sprouts were measured for each of the 54 plants in the test plots.
YEAR 3 (2015)
The third year focused on measuring cumulative impact of alder chip treatments on soils and yields on Peggy Rockefeller farm. Experimental plots locations remained the same and the tomato and Brussels sprout rotation was completed with dry beans (var. Midnight Black Turtle). Field preparation and applications of compost and alder treatments were completed in early June and then direct seeded with on 15 June with an Earthway seeder. All treatments received a Sul-po-mag sidedress and drip tape. Drip tape was installed and mulch then applied one week post-emergence. Plant height and density was recorded in the third week of July and inter-row weed biomass and diversity were sampled in late July and mid-September. Due to uneven senescence patterns between the upper and lower half of the field, final harvests for yield component analysis and yield per row foot began in mid-October and were completed in early November. Soil samples for a soil food web analysis were pulled in 20 September and samples for final soil analyses were pulled on 24 November 2015. Chips remaining from the 2013 harvest also were analyzed for plant available nutrients.
In December, all marked alder “individuals” (clumps of interconnected stumps) were re-measured in terms of number and diameter of new primary and secondary branches sprouted from original base. Reproductive status also was recorded for each individual. Regrowth of the spring chipped alders was not documented.
TOMATO (2013)
There were few significant differences in yields among treatment and between tomato varieties for either Daybreak (DBK) or Peggy Rockefeller (PRF) farms. Initial harvests for DBK (8/13/13) were approximately one week earlier than PRF (8/20/13) and reflect the higher growing-degree-days for Knox County. Final yields per plant for Daybreak Farm (DBK), however, were low due to a short growing season brought on by late blight in mid-August. There were no differences in fruit number nor fruit weight across treatments for either tomato variety. Bellstar, however, tended to have more fruit and slightly higher yields with compost (see Table 1).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
On Peggy Rockefeller Farm, there were no significant a difference in final yields among treatments for Bellstar, although the compost treatment was on average were 5 pounds more than all other treatments (Table 3). The most notable differences in yield on PRF arose after a heat wave in early September. Prior to the heat wave, there were no significant differences in total pounds of fruit across all treatments (Table 2). Post drought, the pounds of OB fruit and number of fruit for the compost treatment was on average 18.7 pounds, nearly 10 pounds greater than the alder amended treatments. This variation of OB tomato yields across treatment was significantly correlated with leaf potassium content (r2=0.89, p<0.001), highest in the compost control (3.3%), intermediate with chip incorporation (2.6%), and lowest in the none, mulch, and spring chip (2.3). (Table 3 and Figure 7)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Table 3 . Post-drought yield of tomatoes on Peggy Rockefeller Farm (10/6/13). avg per plant | |||||
| Orange blossom | Bellstar | ||||
| TREATMENT | number | lb | % K content | number | lb |
| compost | 27.22 | 18.69 | 3.2 | 61 | 20.69 |
| no om | 17.38 | 7.99 | 2.2 | 49.4 | 16.29 |
| alder chip | 17.44 | 10.1 | 2.7 | 42 | 14.67 |
| chip & mulch | 17.33 | 11.36 | 2.5 | 41.6 | 13.51 |
| mulch | 16.67 | 8.58 | 2.3 | 46.67 | 12.23 |
| spring chip | 14.67 | 9.25 | 2.3 | 51.25 | 14.44 |
In conclusion, these results suggest the incorporated winter chips were an important source of potassium in the first growing season and helped ameliorate the stress impact for Orange blossom. Analyses of plant available nutrients from chips a two-year-old chip pile also indicates that alder is a good source of potassium along with all other nutrients except for nitrogen (Table 4).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Figure 7. Variation in tomato stress across treatments
BRUSSELS SPROUTS (2014)
There were no significant treatment effects on Brussels sprouts in 2014. The average weight of Brussels sprout stalk was 5.5 lbs. Sprout weight average across all treatments was 2 pounds per stalk. There appears to be no negative within year effect of the incorporated alder chip and no evidence of nitrogen immobilization (Table 5).
Cabbage white butterfly was a common visitor on the Brussels sprouts after the removal of the Remay in early August. Overall, the percent of the top leaves with some damage ranged from 78 to 95 %, with the highest levels on the compost control and the spring alder chip. However, the extent of damage was more variable, with less than 25 % in the control and increasing across treatments, with the most in the spring alder chip with over 50%. There was no correlation between upper leaf damage and sprout weight.
|
|||||||||||||||||||||||||||||||||||||||||||||
DRY BEANS (2015)
There were no significant differences among treatments in terms of overall yield despite different initial densities (Table 6). Differences in individual size compensated for the initial difference in row density; spring chip plants were larger, had the most pods per plant and highest bean yield (both number and mass) per plant. Overall, drilling into the chipped treatments was awkward and led to uneven densities across treatments. Germination and establishment of plants in the fresh spring chip was lower (by 85%) and slower than the other treatments with final senescence was delayed by over two weeks.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Figure 8. Aerial view of experimental plot
Figure 8. Aerial view of experimental plot

Figure 9. Bean establishment and mean height in July.
2013-2015 trends in SOILS and WEEDS
Soil and weed responses, in contrast, changed over the three years of the experiment. The Chip + mulch treatment received twice the amount of carbon input and there was a noticeable increase in the sifted-fraction of soil organic matter (1.4%) in contrast to the treatment with no soil conditioner with an overall decrease of 3% over the three years of the experiment (Figure 10 and Table 7). The incorporated chip, mulch, and spring chip treatments lost approximately 1% SOM in the sifted-fraction over the same time period. Degree of compaction as measured with a penetrometer, paralleled these patterns of SOM with the highest compaction in the no soil conditioner treatment and lowest across all chip treatments.
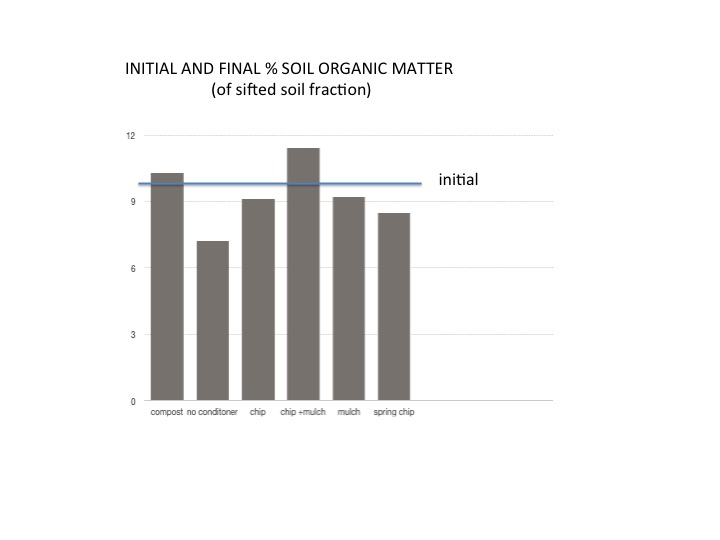 FIGURE 10. Initial and final % soil organic matter across at treatments
FIGURE 10. Initial and final % soil organic matter across at treatments
Soil food web analysis considers the total amounts of bacterial and fungal fractions that are the foundation of soil processes and also measures total active fractions at the time of harvest (Table 8). In general,
Not surprisingly, weed biomass varied with both crop and treatment (Table 9). Overall weed biomass was highest with Brussels sprouts while the beans had consistently lowest. Overall, the mulched treatments had an order of magnitude fewer weeds. The spring chip appeared to have an allelopathic effect, with weed density increasing only modestly with Brussels sprouts and was extremely low in beans. This allelopathic effect also was suggested by the slow and poor germination (85%)of the direct seeded beans.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TABLE 9. Average within-row weed biomass (g) across crop/year and between treatments. | |||||||
| TOMATO | BRUSSELS SPROUT | BEAN | |||||
| TREATMENT | 2013 | 2014 | 2015 | ||||
| compost | 7.6 | 89.4 | 11.0 | ||||
| none | 31.8 | 323.8 | 14.8 | ||||
| chip | 13.4 | 70.7 | 35.8 | ||||
| chip+mulch | 2.4 | 10.7 | 10.0 | ||||
| mulch | 1.5 | 15.0 | 8.3 | ||||
| spring chip | 16.8 | 28.4 | 4.7 | ||||
COPPICING
Alder regrowth was measured for 11 individuals along a 150-foot transect that ran north/south along a hayfield/pasture fence line. The northern half of the transect was shaded by mixed forest to the west and received only morning sun. The southern end of the transect, in contrast, received full sun for the majority of the day.
Of the eleven individuals surveyed, the average cut stump diameter was 65 millimeters, with a range of 41 to 118 mm. The number of primary stump sprouts was similar for each individual along the transect, ranging between 6 to 9 primary stump sprouts. Amount of growth of each primary stump sprout differed significantly, with the diameter of those in the shade on average was 8 mm in on contrast to those to the south of 14 mm (see FIGURE 5).
All of five individuals on the southernmost part of the transect had on average 15 more secondary sprouts associated with the primary sprouts. Three of these five individuals also were in flower in the third year after the initial harvest.
Quote from participating farmer CJ WALKE:
“Alder has been an excellent on-farm resource for mulch to suppress weeds and retain moisture, while slowly building soil organic matter. We have a lot of alder on the farm and cut it back annually to maintain fence lines, so coppicing has been a great method to keep the fence lines clear, while also encouraging regrowth for future harvest. Harvesting alder has been easier in the winter when the ground is frozen, since alder grows in our lowlands, and we can get equipment to the stands compared to spring harvest where the ground is too wet and we have to haul the alder out by hand”.
Education & outreach activities and participation summary
Participation summary:
2013
SUZANNE MORSE
Re-imagining the use of alder as a fertility tree (Eagle Hill Natural History Center, Maine and Norwegian University of Life Sciences)
POLLY McADAM- student researcher
Potential of coppiced alder as an on-farm source of fertility for vegetable production (COA Science Symposium)
Video https://www.youtube.com/watch?v=l1Iu7yB0ADY
CJ WALKE Field presentations to visitors in summer 2013, 2014 and 2015
2014
SUZANNE MORSE
“ Conditioning with woodchips” presentations to Maine Vegetable and Fruit School
Portland (100 registrants)
Bangor (90 registrants)
CJ WALKE
“Using alder chips in vegetable production” at MOFGA farmer-to-farmer conference
Project Outcomes
In terms of spreading out work on a vegetable farm, winter harvest of alder may be most desirable. Time estimate for cutting, chipping, and unloading 2 yards of chip was 8 hours for one person and less than half-a-day for two workers. When necessary, chipper rental costs run about $200 for 24 hours. Arrangements for free alder chips might be available and delivered from road crews.
Areas needing additional study
This study focused on using ramial chips harvested during the New England winter when there is easy access to otherwise wet ground and when farmers might have more time for cutting and stacking branches for chipping. Spring chips have the potential to provide more nitrogen and phosphorous, increased soil microbial activity and the suppression of annual weeds. All of these effects are worthy of further exploration. Overall, the chip + mulch treatment showed a significant increase of SOM over the initial % SOM and suggests that increasing the rate of application from 1 bushel to 2 bushels would further SOM in the sifted fraction for all the other alder treatments in addition to an increase in the bacterial and fungal fractions for the spring chips. Decomposition of spring chip over the summer and application in the spring might decrease potential allelochemical effects on small, direct seeded crops.
Alder is often the focus of various roadside eradication schemes and its use as a companion to farming systems, as a soil conditioner is worthy of consideration for farmers with easy access to alder and a chipper. Loss of Daybreak Farm from this study after the first year of the study (the lower SOM farm) limits how much we can know in how effective or perhaps limiting the addition of this woody material.