Final Report for SW08-052
Project Information
In three certified organic orchards and one research orchard we assessed whether an alfalfa cover crop leads to biological control of pear psylla, an insect pest of pears. We monitored predator and pest densities, predator movement and gut contents of predators in alfalfa and control (grass) plots. Pest or predator densities in the trees were not affected by the cover crop, even though the cover crop was found to produce large numbers of predatory insects, with a proportion of those shown to move from the orchard floor into the tree. Specimens collected for the gut contents analyses are being processed.
1. Determine if an alfalfa cover crop leads to an increase in densities of generalist predators in the pear tree canopy;
2. Examine whether the increased densities of predators in trees having a cover crop understory is due to movement by predators from cover crop to tree;
3. Assess whether those colonists then switch diet to attack pear psylla;
4. Determine if the combined effect of habitat and dietary switching then leads to biological control of psylla and a lowering of damage to fruit;
5. Forward results to growers and evaluate outcomes of the project.
Pear psylla, Cacopsylla pyricola (Förster) (Hemiptera: Psyllidae) is one of the most damaging arthropod pests of pears in North America and Europe (Horton & Unruh 2007). The pest was introduced into the eastern U.S. in the early 1800s from Europe (Slingerland 1892), arriving in western U.S. orchards by the late 1930s (Webster 1939). It now occurs in all pear growing regions of North America. The primary damage caused by pear psylla is fruit russet. Nymphs of pear psylla produce large quantities of honeydew, and as this waste product drips onto fruit, it produces a darkish stain that leads to downgrading of the harvested product and lowered economic return. Currently, $1000/ha or more may be required for its control in pear orchards of California, Oregon and Washington (Horton 1999, Madsen & Dunley 2004), where over 95% of U.S. pears are produced. The primary means of control is with insecticides. Chemical control begins as early as February and may extend well into late summer. These practices prompt concerns about impact of pear pest management on the environment and worker safety. Extensive use of insecticides against psylla has also led to development of resistance in this pest (Follett et al. 1985, Burts et al. 1989, Croft et al. 1989).
The paradigm for pear pest management has recently begun to shift from broad-spectrum chemicals to the use of mating disruption for lepidopteran pests combined with selective chemicals and biological control for pear psylla. This shift in approach has had the effect of making the future for biological control of pear psylla more promising now than it has been since the pest’s arrival in the western U.S. over 60 years ago. Native generalist predators, dominated by the Heteroptera, have long been implicated in psylla control (Madsen et al. 1963, Westigard et al. 1968, Unruh et al. 1995). Other generalist predators, including lacewings, ladybird beetles and spiders may also be locally important (Westigard et al. 1986, Horton et al. 2002).
Cover crops are widely used in fruit or nut orchards to reduce soil erosion, improve soil quality and to help control weed and arthropod pests (Sustainable Agriculture Network 1998). Interest in the use of cover crops in orchards to enhance biological control of pests is shown by the large literature on this topic in technical journals or books (Bugg & Waddington 1994, Pickett & Bugg 1998), in trade journals (Alway 1998) and on web-pages (U.C. Davis 2002). The intent is to use cover crop habitats to supplement natural enemies with resources not provided by the crop, with the expectation that the cover crop will then be a source of natural enemies moving into the tree. The literature on cover crops and biological control in orchards was reviewed in Bugg & Waddington (1994) and includes studies in nut crops (e.g., Bugg et al. 1991, Smith et al. 1996), stone fruits (e.g., Stephens et al. 1998) and pome fruits (e.g., Fye 1983).
Despite the interest shown for using cover crops in orchards, there is little consensus about the actual effects of this technology on biological control. Numerous studies have shown that diversification of agricultural systems (as through cover cropping) leads to higher densities of natural enemies (reviews by Bugg & Waddington 1994, Landis et al. 2000, Langellotto & Denno 2004). However, in those studies it is generally unclear whether the higher densities or diversities actually lead to better pest control (Bugg & Waddington 1994, Denno et al. 2005, Snyder et al. 2005). These uncertainties may be the result of several factors, encompassing both methodological and biological issues. Methodological problems in studies include lack of replication, insufficient plot size, lack of pest suppression data and failure to conduct the research under commercial growing conditions (Bugg & Waddington 1994, Brown & Lightner 1997, Snyder et al. 2005). Limitations in our understanding of natural enemy biology have also contributed to the lack of consensus regarding the value of cover crops. Two limitations are especially prominent and are to be examined in this project:
1. Poor understanding of movement by generalist predators between cover crop and crop habitats; and
2. An incomplete understanding of dietary switching in generalist predators that move from cover crop into the crop.
Cooperators
Research
A. Orchard sites and cover crop
The studies were done in three certified organic orchards and one research orchard. The cover crop in each orchard was alfalfa (Medicago sativa). Alfalfa was planted in two feet wide strips in three organic pear orchards in spring 2008; at the experimental orchard, aisle-wide strips of alfalfa had been planted in spring 2006. Each alfalfa plot was paired with a control (grass) plot. Sampling at the three commercial orchards was initiated in spring 2009; sampling of arthropod densities, predator movement and predator feeding were begun in 2007 at the research orchard. Plot size varied between 0.3 and 0.5 acres per plot at the commercial sites; each plot was 0.2 acres in size at the research orchard.
B. Pest and predator densities
Pear psylla densities were estimated using beat trays to sample adults (20 trays per plot) and leaf samples for eggs and nymphs (200 leaves per plot) following protocols in Horton (1999); samples were taken in both alfalfa and control (grass) plots. Predator numbers were assessed using beating trays (tree, 20 trays per plot), sticky traps (at both canopy- and ground-leve, 10 traps per plot and height) and sweep nets (grass and alfalfa understory, 100 sweeps per plot). Biological control of pear psylla in cover crop vs control plots was assessed by comparing densities of eggs, nymphs and adults of pear psylla between treatment and control plots.
C. Predator movement
Predator movement from orchard understory to tree was quantified by applying a protein marker (liquid egg whites) to the understory and assaying tree-collected predators for presence of the marker using ELISA. The technology was developed by cooperator Vince Jones (Jones et al. 2006) and modified for use in cover crop systems by cooperators on this project (Horton et al. 2009). Liquid egg whites (Organic Valley Farms) were diluted to 10% in water and applied to the cover crop using a boom sprayer attached to an ATV (Fig. 1). We collected arthopods from the tree canopy by jarring tree limbs with a rubber hose and trapping the dislodged predators on a section of butcher’s paper that has been coated with a thin layer of tangletrap (Fig. 2). The arthropods were then removed from the adhesive in the field using wooden toothpicks and placed singly into 1.5 ml microtubes (Fig. 2). Microtubes and specimens were put on ice for transport to the laboratory. Each specimen was washed with 1 ml of tris-buffered saline solution. The wash was then analyzed by indirect ELISA (Jones et al. 2006) for detection of egg proteins. Control arthropods (i.e., known unmarked specimens) for the ELISA were collected off-farm.
D. Predator gut contents
Gut contents were analyzed using molecular methods (Unruh et al. 2008). Field-collected predators were homogenized in microtubes. Each homogenate was tested using ELISA to detect psylla protein. The same predator specimens collected for our marker assays (above) were used to assess gut contents. Thus, once a specimen was washed with buffer to collect egg marker, gut contents of that same specimen was assessed. Control arthropods were collected off-farm in an alfalfa field. Specimens are still being processed at the time of this report.
{kind=link}
{kind=link}
Pest and predator densities
Densities of natural enemies and psylla were very low in all three commercial orchards, in both cover crop and tree canopy (data not shown). Plots were mowed at high frequency in both control and cover crop plots by growers, and we suspect that regular mowing prevented build-up of natural enemies in the cover crop (e.g., Horton et al. 2003). Thus, results presented below are exclusively from the research farm, where mowing was limited to twice per season (spring and early- to mid-summer).
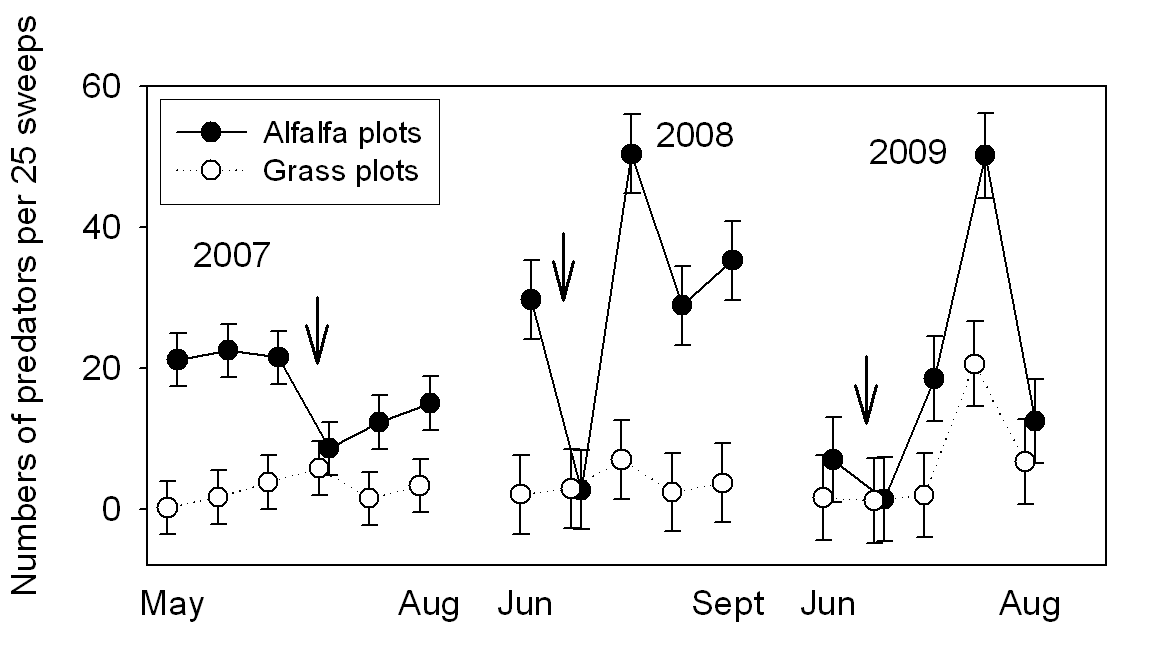
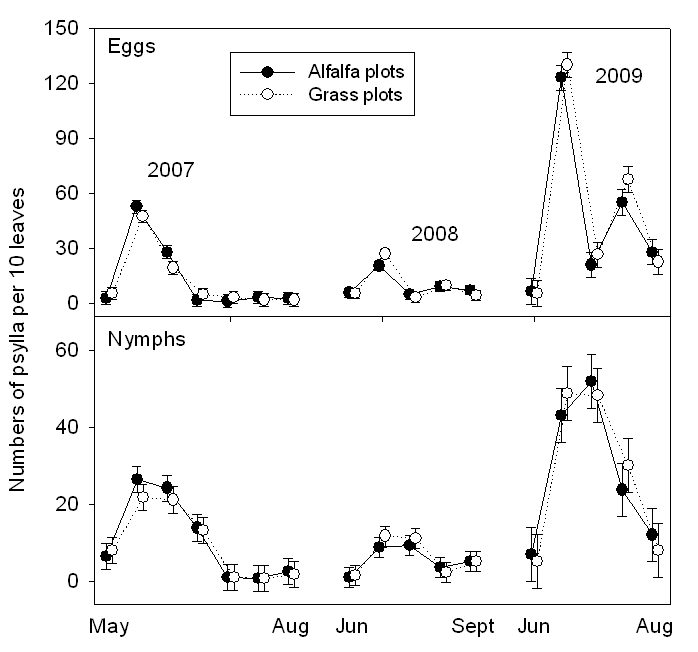
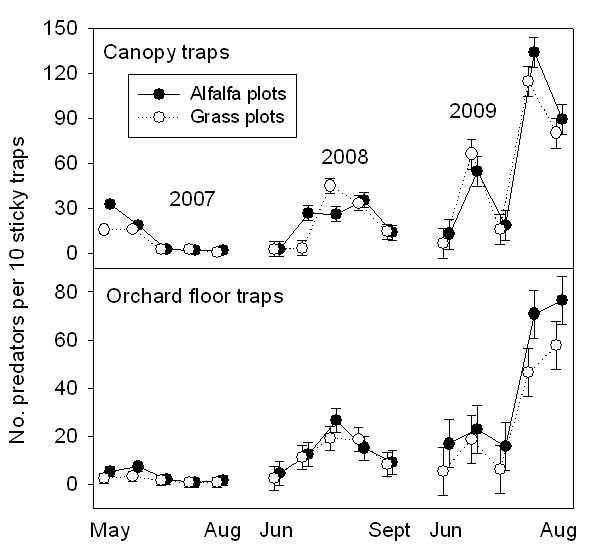
Generalist predators in the tree canopy and orchard understory were dominated by true bugs, ladybird beetles, green lacewings and spiders. Densities of predators in the orchard floor vegetation were several-fold higher in the alfalfa plots than the grass plots (Fig. 3). There was a significant presence all season of predators in the alfalfa cover crop, except immediately following mowing (Fig. 3). Counts of predators in the tree were, if anything, larger in the grass plots than the alfalfa plots (Fig. 4). Sticky trap catches of generalist predators are shown for canopy-height traps (Fig. 5, upper panel) and ground-level traps (Fig. 5, lower panel). Numbers of predators on traps were similar in the alfalfa and grass plots (Fig. 5). Trap catch was dominated by the true bugs and lacewings. Densities of adult psylla were statistically similar in grass and alfalfa plots (Fig. 6). Counts of psylla eggs and nymphs were also similar in grass and alfalfa plots (Fig. 7).
Movement studies
Marker results are shown for the 2009 results, the year for which we have the most complete data (Table 1). Results support the hypothesis that certain predatory taxa moved between orchard floor vegetation and the tree canopy. Lacewings may have been especially mobile. No striking differences were noted between alfalfa and grass plots.
Gut contents analysis
Specimens for the gut contents analyses are still being processed. We show partial results for the 2007 data (Table 2). A large proportion of specimens across taxa showed evidence of having ingested pear psylla, especially nymphs of known psylla predators (Anthocoris spp. and Deraeocoris brevis). A large proportion of adult chrysopids also scored positive for the presence of psylla proteins, even those taxa that do not feed on animal prey in the adult stage (Chrysoperla). Those insects apparently had been feeding upon psylla honeydew (the waste product is detected by this procedure; Unruh unpubl.). We caution that secondary predation (i.e., feeding on an insect which had previously fed upon psylla) could also lead to positive scores, thus results for spiders in particular must be evaluated somewhat cautiously. Ongoing analyses are to include an assessment of whether specimens that carry the marker from the movement study also showed evidence of having fed upon psylla. That analysis will allow us to examine whether specimens that switch habitats will also then switch diet to feed upon the target pest.
Conclusions
We demonstrated that predator densities were much higher in the alfalfa cover crop than the grass understory and showed that some proportion of generalist predators in the tree canopy had originated from (or visited) orchard floor vegetation. Moreover, a large proportion of taxa showed evidence of having fed upon the target pest, pear psylla. Despite these observations, we were unable to show that the alfalfa cover crop translated into higher densities of predators in trees or into reduced densities of pear psylla. There are at least three potential explanations for the lack of effects:
1. Plot sizes (800 m2) were too small to demonstrate effects (i.e., predators that moved from the alfalfa cover crop and into pear trees often colonized trees in nearby control plots, obscuring effects of the cover crop);
2. Predators that were highly abundant in the cover crop and were found to move into the tree canopy failed to attack pear psylla once they had colonized the tree canopy. Our gut contents analyses (ongoing) will allow us to address this hypothesis.
3. The dominant psylla predators in this orchard are known to be Anthocoris spp. and Deraeocoris brevis, which together comprised over 60% of generalist predators obtained on beat trays. However, both predators have a strong preference for arboreal habitats and were found only irregularly in the samples taken from alfalfa. Thus, densities of these psylla predators in trees are almost certainly independent of what plant species are used as a ground cover.
Alway, T. 1998. Can cover crops improve biological control of pests? Good Fruit Grower (Aug. 1998): 55-56.
Brown, M.W. and G.W. Lightner. 1997. Recommendations on minimum experimental plot size and succession of aphidophaga in West Virgina, USA, apple orchards. Entomophaga 42: 257-267.
Bugg, R.L. and C. Waddington. 1994. Using cover crops to manage arthropod pests of orchards: a review. Agric. Ecosyst. Environ. 50: 11-28.
Bugg, R.L., J.D. Dutcher & P.J. McNeill. 1991. Cool-season cover crops in the pecan orchard understory: effects on Coccinellidae (Coleoptera) and pecan aphids (Homoptera: Aphididae). Biol. Control 1: 8-15.
Burts, E.C., H.E. van de Baan, and B.A. Croft. 1989. Pyrethroid resistance in pear psylla, Psylla pyricola Foerster (Homoptera: Psyllidae), and synergism of pyrethroids with piperonyl butoxide. Can. Entomol. 121: 219-223.
Croft, B.A., E.C. Burts, H.E. van de Baan, P.H. Westigard, and H. Riedl. 1989. Local and regional resistance to fenvalerate in Psylla pyricola Foerster (Homoptera: Psyllidae) in western North America. Can. Entomol. 121: 121-129.
Denno, R.F., D.L. Finke and G.A. Langellotto. 2005. Direct and indirect effects of vegetation structure and habitat complexity on predator-prey and predator-predator interactions. In P. Barbosa and I. Castellanos (eds), Ecology of predator-prey interactions. Oxford.
Follett, P.A., B.A. Croft, and P.H. Westigard. 1985. Regional resistance to pesticides in Psylla pyricola from Oregon pear orchards. Can. Entomol. 117: 565-573.
Fye, R.E. 1983. Cover crop manipulation for building pear psylla (Homoptera: Psyllidae) predator populations in pear orchards. J. Econ. Entomol. 76: 306-310.
Horton, D.R. 1999. Monitoring of pear psylla for pest management decisions and research. Integrated Pest Mgmt. Rev. 4: 1-20.
Horton, D.R., D.A. Broers, T. Hinojosa, T.M. Lewis, E.R. Miliczky and R.R. Lewis. 2002. Diversity and phenology of predatory arthropods overwintering in cardboard bands placed in pear and apple orchards of central Washington state. Ann. Entomol. Soc. Am. 95: 469-480.
Horton, D.R., D.A. Broers, R.R. Lewis, D. Granatstein, R.S. Zack, T.R. Unruh, A.R. Moldenke and J.J. Brown. 2003. Effects of mowing frequency on densities of natural enemies in three Pacific Northwest orchards. Entomol. Exp. Appl. 106: 135-145.
Horton, D.R. and T.R. Unruh. 2007. Pear insects: ecology and control. In D. Pimentel (ed), Encyclopedia of pest management. Volume II: 474-477. CRC Press, NY.
Horton, D.R., V.P. Jones and T.R. Unruh. 2009. Use of a new immunomarking method to assess movement by generalist predators between a cover crop and tree canopy in a pear orchard. Amer. Entomol. 55(#1): 49-56.
Jones, V. P., Hagler, J. R., J. F. Brunner, C. C. Baker, T. D. Wilburn. 2006. An inexpensive immunomarking technique for studying movement patterns of naturally occurring insect populations. Environ. Entomol. 35: 827-836.
Landis, D.A., S.D. Wratten and G.M. Gurr. 2000. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 45: 175-201.
Langellotto, G.A. and R.F. Denno. 2004. Responses of invertebrate natural enemies to complex-structured habitats: a meta-analytical synthesis. Oecologia 139: 1-10.
Madsen, H.F., P.H. Westigard and R.L. Sisson. 1963. Observations on the natural control of the pear psylla, Psylla pyricola Förster, in California. Can. Entomol. 95: 837-844.
Madsen, T.M. and J.E. Dunley. 2004. Areawide organic pest management. 100th Ann. Meeting of the Washington State Horticultural Assoc., Yakima, WA.
Pickett, C.H. and R.L. Bugg (eds). 1998. Enhancing biological control: habitat management to promote natural enemies of agricultural pests. Univ. Calif., Berkeley.
Slingerland, M.V. 1892. The pear-tree psylla. Vol. 44, pp. 161-186, Ithaca, NY: Cornell Univ. Agric. Exp. Station Bull.
Smith, M.W., D.C. Arnold, R.D. Eikenbary, N.R. Rice, A. Shiferaw, B.S. Cleary and B.L. Carroll. 1996. Influence of ground cover on beneficial arthropods in pecan. Biol. Control 6: 164-176.
Snyder, W.E., G.C. Chang and R.P. Prasad. 2005. Conservation biological control: biodiversity influences the effectiveness of predators. In P. Barbosa and I. Castellanos (eds), Ecology of predator-prey interactions. Oxford.
Stephens, M.J., C.M. France, S.D. Wratten and C. Frampton. 1998. Enhancing biological control of leafrollers (Lepidoptera: Tortricidae) by sowing buckwheat (Fagopyrum esculentum) in an orchard. Biocontrol Sci. Tech. 8: 547-558.
Sustainable Agriculture Network. 1998. Managing cover crops profitably. Sustainable Agriculture Network, Washington, D.C.
U.C. Davis. 2002. http://www.sarep.ucdavis.edu
Unruh, T.R., P.H. Westigard and K.S. Hagen. 1995. Pear psylla. In J.R. Nechols, L. Andres, J.W. Beardsley, R.D. Goeden and G. Jackson (eds.), Biological control in the western United States. DANR Publications, Univ. California at Davis.
Unruh, T.R., T. Yu, L.S. Willett, S.F. Garczynski and D.R. Horton. 2008. Development of monoclonal antibodies to pear psylla, Cacopsylla pyricola (Hemiptera: Psyllidae) and evaluation of field predation by two key predators. Ann. Entomol. Soc. Am. 101: 887-898.
Webster, R.L. 1939. The pear psylla survey. Proc. Wash. State Hort. Assoc. 35: 36-40.
Westigard, P.H., L.G. Gentner and D.W. Berry. 1968. Present status of biological control of pear psylla in southern Oregon. J. Econ. Entomol. 61: 740-743.
Westigard, P.H., L.J. Gut and W.J. Liss. 1986. Selective control program for the pear pest complex in southern Oregon. J. Econ. Entomol. 79: 250-257.
- Figure 3. Numbers of predators per 25 sweeps.
- Figure 6. Numbers of adult pear psylla per 20 trays.
- Table 2. Mean percentage (averaged over plot) of predator specimens having pear psylla protein in gut.
- Figure 7. Numbers of immature psylla per 10 leaves.
- Table 1. Number of marked/total (% marked) predators.
- Figure 4. Numbers of predators per 20 beat trays.
- Figure 5. Numbers of predators per 10 sticky traps.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
This is the first study to demonstrate empirically that generalist predators move between cover crop and tree canopy in an orchard situation. Once the gut contents analyses have been finished, we will also have information showing that predators which switch habitats are also generalized enough to switch diet to feed upon the target pest.
Research outcomes
Education and Outreach
Participation summary:
This project was discussed at a cover crop field day (sponsored by Washington State University Extension), located at an apple orchard near Quincy, WA (August 2009; attendance approximately 20 people). A slide presentation showing results of this project was made at an Organic Farming workshop in Ellensburg, WA (March 2010; sponsored by Washington State University; attendance approximately 30 people). Results of the project were also presented at the 2008 and 2009 annual Northwest Pear Research Reviews (Yakima and Wenatchee, WA). These meetings are attended by large numbers of pear growers from Oregon and Washington.
Publication:
Horton, D.R., V.P. Jones, and T.R. Unruh. Assessing habitat and dietary switching by generalist predators in an orchard – cover crop system. Biological Control (in preparation; manuscript to be completed winter 2010-11).
Education and Outreach Outcomes
Areas needing additional study
It would be useful to know whether a cover crop species less vigorous than alfalfa might be more compatible than alfalfa with practices used by commercial growers in managing orchard floor vegetation.