2008 Annual Report for GNC07-079
Effects of two Conservation Reserve Program Prairie Restorations on Soil Microbial Communities
Summary
The federal Conservation Reserve Program (CRP) provides payments to agricultural producers who take highly erodible or marginal land out of production and restore natural habitats such as forest or prairie. However, societal incentives to producers to grow biomass for use as biofuels may compete with traditional set-aside programs like the CRP, potentially impacting the soil quality gains made by the CRP over the last twenty years.
This study aims to investigate soil biological quality and soil carbon under CRP prairie restorations compared to maize cultivation.
Objectives/Performance Targets
- Evaluate the effects of CRP management on microbial biomass, soil microbial community composition and diversity. Relate microbial indices to soil quality parameters (especially soil carbon) that may respond to CRP and post-CRP cultivation practices. Quantify soil carbon differences between paired CRP and cultivated sites. Examine the relationship between soil aggregates and soil carbon within CRP and cultivated sites. Increase farmer knowledge of soil quality, and increase the ability of farmers to preserve long-term soil quality through dissemination of educational materials and decision-making tools.
Accomplishments/Milestones
Study Sites and Sampling Procedure
Samples were collected in 2008 from two private farms in southwestern Dane County (designated Site A and Site B). The sites were within 5 km of each other to ensure similar climatic conditions. Both sites contained “treatment” plots of mature CRP prairie restoration paired in space with treatment plots of corn cultivation. At Site A, the prairie restoration was 5 years old and located on a north-facing slope. Management of the prairie included a controlled burn in spring 2008. The plot of corn cultivation at Site A was in a 2-year rotation with soybeans for 10 years, and had not been tilled during this time. At Site B, the CRP prairie restoration was over 10 years old, while the plot of corn cultivation was in its first year of cultivation after having previously been part of the CRP restoration. The corn had been planted under no-tillage conditions and was preceded in the season by tilled winter wheat.
Sampling was conducted during one week in August 2008, following plant peak growth. At each plot (one prairie and one cornfield at each site), we sampled an area of approximately 5200 square meters (Figure 1). Fifteen soil samples per plot were taken along a 450 meter transect, each sample made up of five composite cores in order to reduce sampling variability. Each core for compositing was taken with a push probe of 5 cm diameter to a depth of 25 cm. Cores were broken into two depths, 0-10 cm, and 10-25 cm. We also took ten bulk density cores per plot, along the same transect.
Following sampling, soils were transported immediately to the laboratory, where we removed large roots (> 4 mm) and rocks, and split the samples into two subsamples. We homogenized and froze one set of subsamples at -20º C for microbial analysis, and another subset was kept in plastic bags in a field-moist state until physical fractionation one week later.
Physical and Chemical Analyses of Bulk Soils
We analyzed each sample for total carbon and total nitrogen analysis using a Elementar VarioMax CN analyzer (Elementar, Hanan, Germany). We measured soil pH by 1:1 determination of air-dried soil in deionized water (Thomas 1996). We measured soil texture by particle size analysis by the hydrometer method. We obtained soil water content gravimetrically by drying soil at 110º for 24 hours and quantifying water loss.
In 2009, we analyzed each sample for total carbon and total nitrogen analysis using a Elementar VarioMax CN analyzer (Elementar, Hanan, Germany). We measured soil pH by 1:1 determination of air-dried soil in deionized water (Thomas 1996). We measured soil texture by particle size analysis by the hydrometer method. We obtained soil water content gravimetrically by drying soil at 110 degrees Celsius for 24 hours and uantifying water loss.
We measured soil microbial community composition from bulk soils in each treatment using membrane lipid analyses, as described by Smithwick et al. (2005). Since phospholipids in cell membranes are metabolized rapidly upon cell death, phospholipid analysis of soil microorganisms provides an estimate of viable microbial biomass White et al., 1979). Briefly, we extracted, purified, and identified cell membrane lipids using a PLFA extraction based upon a modified Blig and Dyer (1959)technique, followed by modified FAME analysis (Microbial ID Inc., Hayward, CA, USA).
To better assess the relationships between microbial community composition and soil structure among different treatments, we chose a subset of samples from each treatment to fractionate into different sized soil aggregates. Aggregates were separated by size according to a modified wet-sieving procedure (Elliott 1986), also described in Simpson et al (2004). We performed a physical fractionation by wet-sieving of 24 fresh samples (six per plot), into macroaggregates (>250 um), microaggregates (53-250 um), and the silt and clay fraction (<250 um).
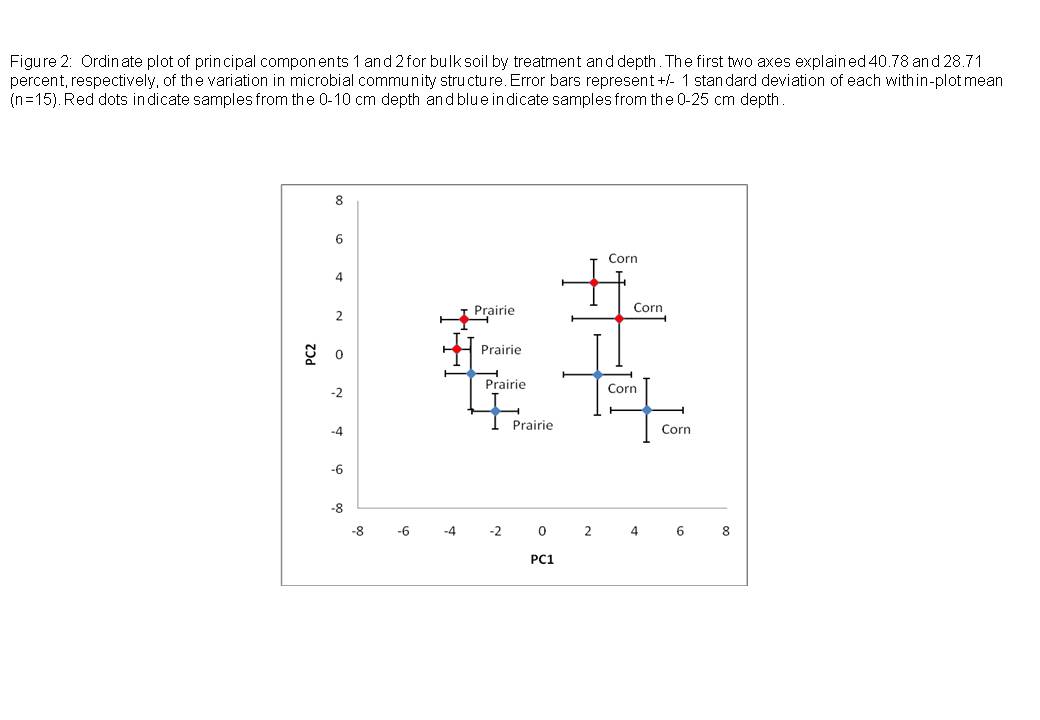
We used principal component analysis on the arcsin transformed mol fraction (relative abundance) of lipids to provide a general microbial lipid ‘fingerprint’ (JMP, SAS Institute, 2002) of each treatment and depth sampled. An independent sample t-test on the principal components (PC1 and PC2) was conducted to assess the effects of treatment on overall microbial community structure. To assess possible treatment effects on physico-chemical response variables, and those of total microbial biomass, individual lipid biomass and relative abundance, microbial guild biomass and relative abundance), we used unpaired t-tests of the means of prairie and corn sites (n=2). To assess differences in depth on these response variables, we used paired t-tests between the 0-10 cm and 10-25 cm depths of all treatments (n=4). Table 2 summarizes the statistical analyses we utilized.
We also tested the effects of soil fraction on microbial and physical variables (Table 2). We used two-way analysis of variance(ANOVA) to test the effects of treatment by soil fraction for total carbon and total nitrogen, and an unpaired t-test of the effects of treatment on % macroaggregates, % microaggregates, and % silt and clay fraction. For tests of the effect of soil fraction on microbial community structure, we also used principal component analysis on the arcsin transformed mol fraction (relative abundance) of lipids.
We conducted two-way ANOVA on the principal components PC1 and PC2 to determine the effects of soil fraction and land-use treatment on overall microbial community structure. All ANOVA tests were followed by Tukey’s post-hoc analysis for pairwise comparisons (JMP, SAS Institute 2002).
Our results indicate that CRP prairie restorations can substantially affect soil microbial community properties. Overall microbial community structure differed substantially between prairie restorations and adjacent corn plots. This was the case for both Site A and Site B, despite significant differences in land-use history between the two sites; Site B had only been removed from the CRP restoration and planted to corn for one year, whereas both the treatments at Site A had been planted for a number of years.
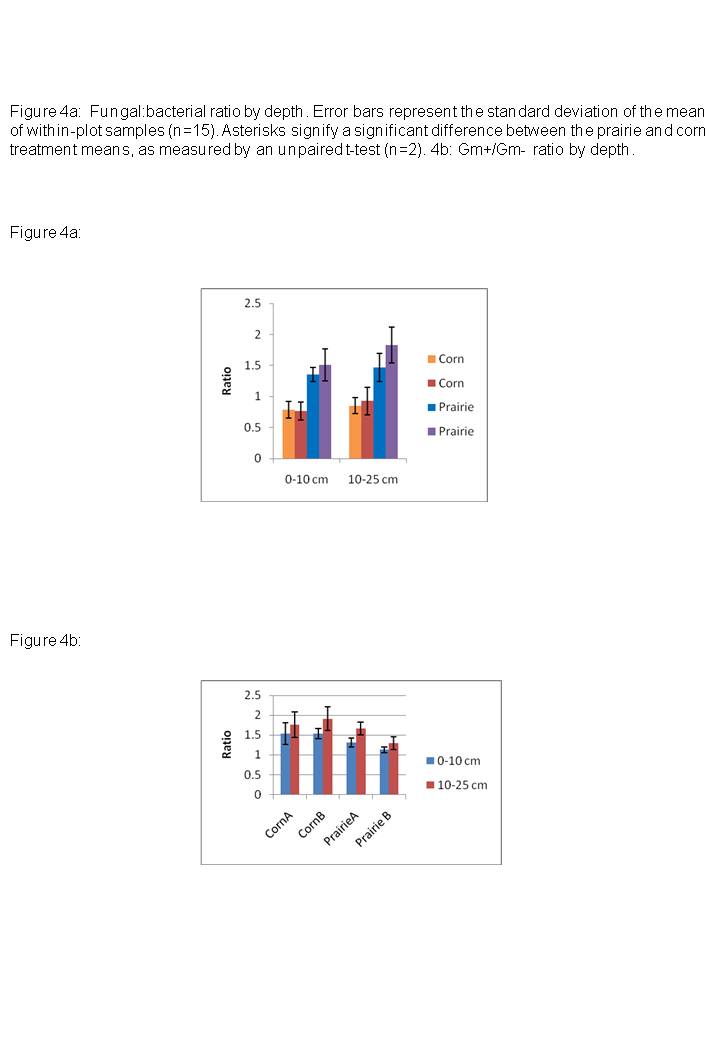
Additionally, the relative abundances of important microbial functional guilds, such as arbuscular mycorrhizal fungi, shifted dramatically between prairies and corn plots, suggesting that prairie restorations can be effective in changing soil biological properties even though basic physico-chemical properties of the sites remained unaffected. We suggest that the total microbial biomass, the fungal:bacterial ratio, and the proportion of AM fungi, are particularly important indicators to measure in determining soil quality in ex-arable or reclaimed land. This is in line with calls
from other restoration ecologists (Harris 2009).
We found a trend toward higher overall microbial biomass within CRP plots compared to the no-till corn plots. This concurs with previous research; for example, Staben et al. (1997) found that CRP prairie restorations increase the total biomass and microbial activity of the soil. Since microbial biomass generally correlates well with carbon inputs into the soil system, it is likely that greater microbial biomass in CRP restorations reflects the effects of dense, perennial vegetation providing greater aboveground litter. Prairie systems are also more likely than corn systems to have dense, fine root systems, thus likely exuding greater amounts of carbon substrate to the soil through the root, and elevating the total microbial biomass by boosting populations in the rhizosphere.
In conclusion, we found significant differences between soil microbial communities within CRP prairie restorations and adjacent reduced-tillage cornfields. In particular, microbial communities could be differentiated by the fungal:bacterial ratio, and the proportion of Gm+ and AM fungi, as well as elevated cyclopropyl/precursor ratios in corn plots. These may be particularly important indicators to measure in determining effects of land-use on soil quality. As a next step, the functional role of these microbial groups in processes important to soil quality, such as carbon dynamics, nutrient mineralization, and soil aggregation, should be further investigated.
- Statistical Analysis Used
- Fungal:bacterial and Gram+:Gram- Ratios
- Microbial Guilds by Treatment
- Ordinate Plot of Microbial Community Structure
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Impacts and Contributions/Outcomes
Many farmers in Wisconsin may be facing financial incentives to remove land from the CRP. Our results suggest that even if basic soil test parameters (such as pH, total carbon, and nitrogen) are not demonstrably changed by conversion of CRP land to corn cropland, soil microbial community composition may be affected, especially the ratio of AM fungi to bacterial biomass. Shifts in the abundance or dominance of AM fungi may have important consequences for soil quality. They have been shown to contribute substantially to the formation of macroaggregates in soils by binding soil particles together with fungal hyphae, and by production of the binding agent glomalin (Rillig et al. 2002, Hallett et al. 2009). AM fungi can also contribute substantially to plant productivity, especially in infertile soils, by increasing plant acquisition of soil nutrients
Wilson and Hartnett 1997).
Our results suggest that no-till management of corn may not significantly alter physico-chemical measures of soil quality enough to be detrimental to soil functions. This is in contrast to conventionally tilled soils, which have been found to have lower soil organic carbon as well as lower measures of soil aggregation (Denef et al. 2004). The lack of difference in soil physico-chemical parameters between corn and prairie treatments does not necessarily suggest equivalence of treatments on soil quality, because of underlying soil and landscape factors that might also be influencing the soil habitat. Many studies register gains in soil quality through participation in the CRP, and our results are not incompatible with those findings. However, several recent studies have cast doubt on the conventional wisdom that soil quality, and soil carbon sequestration, is necessarily improved by CRP prairie restoration (Brye et al. 2002, Kucharik 2007). Effects of CRP management, compared to cropland on the similar soil type, may be strongly affected by plant species composition and management practices such as burning or control of invasive and woody species (Kucharik 2007).
On a similar note, restored prairies, even decades after restoration, have been found not to have recovered essential ecosystem functions and services that native prairies provide, and soil microbial community profiles remain distinct (McKinley et al. 2005). These cautionary studies suggest that microbial community composition, as well, may display variable responses to CRP restorations. This may be especially the case if future CRP prairies are managed in diverse ways, for instance, if prairie plantings are harvested for cellulosic biomass, or fertilized to boost aboveground yield.
Future managers of perennial systems, whether they include prairie restorations for aesthetic purposes, governmental incentives, biomass production for harvest, or carbon sequestration, should be aware that yearly follow-up management of restorations to specifically favor soil quality may be necessary to achieve positive environmental benefits.
Collaborators:
graduate student
University of Wisconsin
3 Battery Bend Ct.
Montgomery Village, MD 20886
Office Phone: 3019901035