Summary:
Inhibitory effects of mustard cultivars, Brassica juncea L. ‘Florida ‘Broadleaf’ and Sinapis alba L. ‘Tilney’ on Phytophthora capsici were tested in the laboratory, greenhouse and four field experiments. In the laboratory experiments, inhibitory effects of 0.22 µm-filtered extracts of FBL, Tilney, a 50:50 mixture of FBL+ Tilney, and control (sterile-distilled water) treatments were evaluated on four P. capsici isolates (PC-1, PC-2, PC-3, and PC-4). The FBL extract significantly reduced colony growth of all P. capsici isolates and sporangia production of PC-1 isolate compared to that of Tilney extract and control treatment. None of the mustard extracts significantly inhibited zoospore germination of P. capsici isolates. The FBL extract significantly reduced P. capsici oospore germination compared to that of control treatment. In the greenhouse trial, extracts of FBL, Tilney, a 50:50 mixture of FBL+Tilney and control (sterile-distilled water) treatments were tested on P. capsici crown infection of ‘Eureka’ cucumber, ‘Magic Lantern’ pumpkin, and ‘Dickinson’ pumpkin seedlings. P. capsici infection on ‘Dickinson’ pumpkin was significantly reduced in presence of FBL + Tilney and FBL extracts compared to Tilney extract and control treatment. In the four field trials (2008-spring, 2008-fall, 2009-spring, and 2010-spring), mean glucosinolate content in mustard plants incorporated to 1 sq. m area of the plots was the lowest in 2008-fall and the highest in 2008-spring trials. P. capsici infection of ‘Eureka’ cucumber, ‘Magic Lantern’ pumpkin, and ‘Dickinson’ pumpkin were assessed in the plots incorporated with FBL, Tilney, a 50:50 seed mixture FBL+Tilney, and control (no mustard) treatments. None of the mustard treatments significantly reduced vine- and fruit-infection by P. capsici in the cucurbit crops tested.
Introduction:
Phytophthora capsici has become a serious threat to cucurbit production in the United States (US) and worldwide in recent years (Babadoost and Zitter, 2009; Hausbeck and Lamour, 2004; Islam, et al. 2004). This pathogen can infect cucurbit plants at any growth stage causing up to 100% crop loss (Babadoost and Islam, 2003). P. capsici oospores can survive in the soil for more than three years (Babadoost and Pavon, 2009). The oospore survival in soil and subsequent germination of oospore is the biggest hurdle for management of P. capsici in the soil. There is no cucurbit cultivar available with measurable resistance to P. capsici (Babadoost and Islam, 2003; Gevens et al. 2006). Further, no single method provides adequate protection against this pathogen (Babadoost and Islam, 2003; Hausbeck and Lamour, 2004; Hwang and Kim, 1995). Pathogen exclusion, crop rotation, moisture management, and fungicide application have been used to reduce the severity of P. capsici infection in cucurbits (Hausbeck and Lamour, 2004). However, none of these methods provides complete control of P. capsici infection in the field (Babadoost and Islam, 2003).
Biofumigation using Brassica crops could be used as a viable alternative to manage P. capsici infection of cucurbits. Biofumigation refers to the release of toxic gaseous materials from wounded tissues of Brassicaceae plant family (Kirkegaard et al., 1993). Wounded Brassica tissues release glucosinolates, which undergo hydrolysis in presence of the enzyme myrosinase to form nitriles, isothiocyanates, and thiocyanates (Cole, 1976; Delaquis and Sholberg, 1997; Rosa et al., 1997). The hydrolysis products of glucosinolates from Brassica crops were reported to have fungicidal activities (Brown and Morra, 2005). Brassica juncea and Sinapis. alba suppressed the germination of Pythium deliense Meurs and P. ultimum Trow. var. ultimum in soil samples (Lazzeri and Manici, 2001). B. juncea has been reported to inhibit the hyphal growth of P. ultimum and Rhizoctonia solani (Snapp et al., 2007). Manici et al. (1997) reported that glucosinolates isolated from B. juncea and S. alba when mixed with myrosinase from S. alba, were toxic to fungi including Fusarium culmorum, F. oxysporum and R. solani. Dunne et al. (2003) reported that B. juncea effectively suppressed the growth of Phytophthora cactorum, and Phytophthora cinnamomi. In this study, effectiveness of B. juncea L. ‘Florida Broadleaf’ and S. alba L. ‘Tilney’ were tested on Phytophthora capsici in the laboratory, greenhouse and field experiments.
REFERENCES
Babadoost, M. and S.Z. Islam. 2002. Phytophthora blight on pumpkin. Plant Health Progress doi:10.1094/PHP-2002-1216-01-DG.
Babadoost, M. and S.Z. Islam. 2003. Fungicide seed treatment effects on seedling damping-off of pumpkin caused by Phytophthora capsici. Plant Dis. 87:63-68.
Babadoost, M. and C. Pavon. 2009. Survival of Phytophthora capsici in soil. In: Proceedings of the 2nd International Phytophthora capsici conference. (p. 2). Duck Cay, Florida.
Babadoost, M. and T.A. Zitter. 2009. Fruit rots of pumpkin: a serious threat to the pumpkin industry. Plant Dis. 93:772-782. doi: 10.1094/PDIS-93-8-0772.
Brown, J. and M.J. Morra. 2005. Glucosinolate-containing seed meal as a soil amendment to control plant pests. NREL/SR-510-35254. University of Idaho, Idaho, USA.
Cole, R.A. 1976. Isothiocyanates, nitriles and thiocyanates as products of autolysis of glucosinolates in Cruciferae. Phytochemistry 15:759-762.
Delaquis, P.J. and P.L. Sholberg. 1997. Antimicrobial activity of gaseous allyl Isothiocyanate. J. Food Prot. 60:943-947.
Dunne, C.P., B. Dell, and G.E. St. J. Hardy. 2003. The effect of biofumigants on the vegetative growth of five Phytophthora species in vitro. Acta Hort 602: 45-51.
Fan, C.M., G.R. Xiong, P. Qi, G.H. Ji, and Y.Q. He. 2008. Potential biofumigation effects of Brassica oleracea var. caulorapa on growth of fungi. Phytopathology 156:321-325.
Gevens, A.J., K. Ando, K.H. Lamour, R. Grumet, and M.K. Hausbeck. 2006. A detached cucumber fruit method to screen for resistance to Phytophthora capsici and effect of fruit age on susceptibility to infection. Plant Dis. 90:1276-1282.
Hausbeck, M.K. and K.H. Lamour. 2004. Phytophthora capsici on vegetable crops: Research progress and management challenges. Plant Dis. 88:1292-1303.
Hwang, B.K. and C.H. Kim. 1995. Phytophthora blight of pepper and its control in Korea. Plant Dis. 79:221-227.
Islam, S.Z., M. Babadoost, K.N. Lambert, A. Ndeme, and H.M. Fouly. 2004. Characterization of Phytophthora capsici isolates from processing pumpkin in Illinois. Plant Dis. 89:191-197.
Kirkegaard, J.A., P.A. Gardner, J.M. Desmarchelier, and J.F. Angus. 1993. Biofumigation – using Brassica species to control pests and diseases in horticulture and agriculture, p. 77-82. In: N. Wratten and R. J. Mailer (Eds.), Proceedings of the 9th Australian assembly on Brassicas. Agricultural Research Institute, Wagga Wagga.
Kushad, M.M., R. Cloyd, and M. Babadoost. 2004. Distribution of glucosinolates in ornamental cabbage and kale cultivars. Sci. Hort. 101:215-221.
Larkin, R.P. and T.S. Griffin. 2007. Control of soilborne potato diseases with Brassica green manures. Crop Prot. 26:1067-1077.
Lazzeri, L. and L.M. Manici. 2001. Allelopathic effect of glucosinolate-containing plant green manure on Pythium sp. and total fungal population in soil. Hort. Sci. 36:1283-1289.
Manici, L.M., L. Lazzeri, and S. Palmieri. 1997. In vitro fungitoxic activity of some glucosinolates and their enzyme-derived products toward plant pathogenic fungi. J. Agric. Food Chem.45:2768-2773.
Miller, P., W. Lanier, and S. Brandt. 2001. Using growing degree days to predict plant stages.
Montguide MT 200103 AG 7/2001. Montanta State University Extension Service, Bozeman, Montana.
Pavon, C.F., M. Babadoost, and K.N. Lambert. 2008. Quantification of Phytophthora capsici oospores in soil by sieving-centrifugation and real-time polymerase chain reaction. Plant Dis. 92:143-149.
Rosa, E.A.S., R.K. Heaney, G.R. Fenwick, and C.A.M. Portas. 1997. Glucosinolates in crop plants. Hort. Rev. 19:99-215.
Snapp, S.S., K.U. Date, W. Kirk, K. O’Neil, A. Kremen, and G. Bird. 2007. Root, shoot tissues of Brassica juncea L. cv. Florida Broadleaf (FBL) and Cereal secale promote potato health. Plant and Soil 294:55-72.
Tian, D. and M. Babadoost. 2004. Host range of Phytophthora capsici from pumpkin and pathogenicity of isolates. Plant Dis. 88:485-489.
Project Objectives:
This proposal was a two-year project to determine the effectiveness of mustard short-cycle cover crops in managing soil-borne Phytophthora capsici and Fusarium spp in cucurbit fields. The specific objectives are: (i) to compare the effects of the planting seasons of mustard on glucosinolate profile and their release in to the soil; (ii) to determine the specific glucosinolates and their concentration in leaves, root and stem; and (iii) to determine the relationship between allelochemical composition, concentration and pest suppression of the chemicals with fungicidal activities for developing biocontrol integrated pest management (IPM) strategies.
Materials and methods:
Laboratory experiments.
FBL, Tilney, and a 50:50 combination of FBL+Tilney were the treatments tested to determine their inhibitory effect on colony growth, sporangial production, zoospore germination, and oospore germination of four P. capsici isolates (PC-1, PC-2, PC-3, and PC-4) in the laboratory. Sterile distilled water (SDW) (control) was included as the control check. Mustard plant tissues were ground in sterile-distilled water (SDW) using a Waring commercial laboratory blender (Waring Products Division, Conair Corp., McConnellsburg, PA). The extract was then passed through eight layers of cheese-cloth. The extracts were immediately filtered through 0.22 µm Millex syringe-driven filter unit (Millipore Corp., Billerica, MA) into sterilized beakers.
Mustard extracts made from fifty grams of mustard plant tissues as described above was used for testing colony growth of the four P. capsici isolates (PC-1, PC-2, PC-3, and PC-4). Ten milliliters of 0.22 µm filtered mustard extract from each of FBL, Tilney, and FBL+Tilney mustards and sterile distilled water (SDW; control) were pipetted separately into 8 cm-diameter Petri plates. A 6-mm plug of P. capsici colony growing on lima bean agar (LBA) was cut and placed on LBA in another Petri plate. The plate with P. capsici plug was inverted onto the plate containing the mustard extract and both plates were taped together with parafilm. This procedure was repeated for all of the P. capsici isolates (PC-1, PC-2, PC-3, and PC-4), mustard extracts (FBL, Tilney, and FBL+Tilney) and control treatment (SDW). The Petri plates were incubated at 24ºC under 12 hr light/12 hr darkness. Radii of the colonies were measured after 5 days. In addition, number of P. capsici sporangia produced was also quantified after five days from the same Petri plates using a light microscope. The counting was done at four different fields of view of a SZX12 microscope (Olympus Corp., Tokyo, Japan) on each plate. This experiment was designed as a factorial experiment in completely randomized design (CRD) with four replications. The experiment as a whole was repeated four times.
The same procedure was also carried out for Fusarium isolates (F-1, F-2, F-3, and F-4) with the same mustard extracts (FBL, Tilney, and FBL+Tilney) and control treatment (SDW). All of the Petri plates were incubated at 24ºC under 12 hr light/12 hr darkness. Diameters of the colonies were measured after 5 days. This experiment was designed as a factorial experiment in completely randomized design (CRD) with four replications.
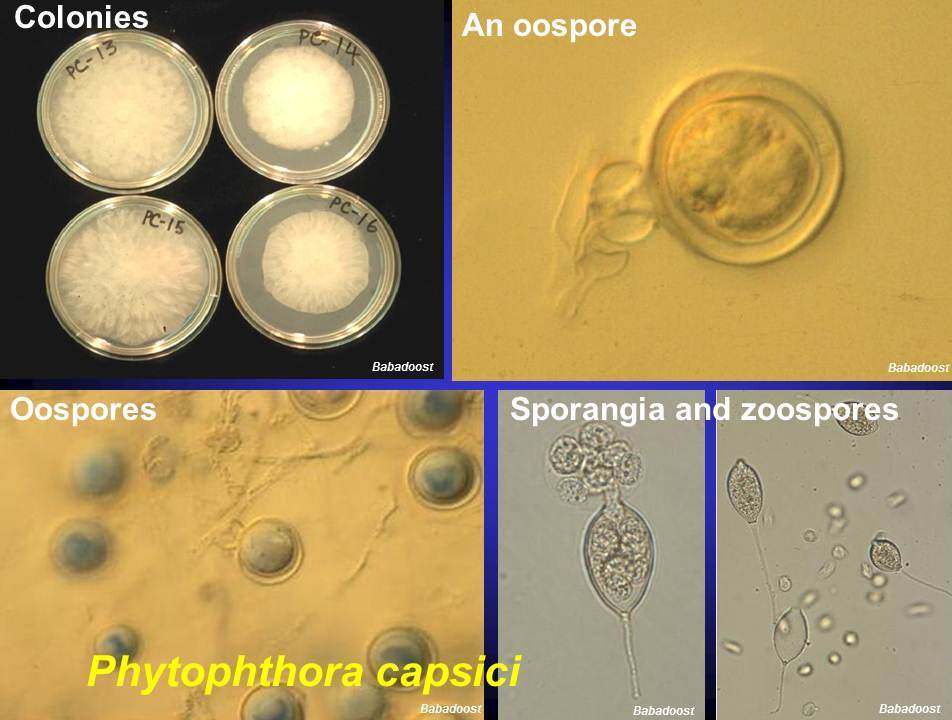
Zoospores of P. capsici isolates PC-1, PC-2, PC-3, and PC-4 were produced using the method by Babadoost and Islam (2002). Two hundred microliters of zoospore suspension in SDW (1,000 zoospores/ml) was added onto each lima bean agar (LBA) plate. Extracts of FBL, Tilney, FBL+Tilney, and SDW (control) were prepared as previously described. Ten milliliters of each of FBL, Tilney, and FBL+Tilney and SDW was pipetted into different Petri plates. The Petri plate with zoospores was inverted over the Petri plate containing the mustard extract and both plates were taped together using parafilm. All of the Petri plates were incubated at 24ºC. Number of germinated zoospores were determined by using SZX12 microscope (Olympus Corp., Tokyo, Japan). This experiment was performed as a factorial experiment in CRD with four replications. The experiment as a whole was repeated four times.
P. capsici oospores were tested for germination in presence of 0.22 µm-filtered extracts of FBL, Tilney, and FBL+Tilney and SDW (control) treatments. P. capsici oospores were produced by pairing up isolates of PC-1, PC-2, and PC-3 (A1 mating type) with PC-4 (A2 mating type) using the method described by Pavon et al. (2008). Two hundred microliters of oospore suspension in SDW (500 oospores/ml) was added into sterilized 1.5 ml microfuge tube containing 800 µl of 0.22 µm-filtered mustard extract. All of the microfuge tubes were incubated at 24ºC in darkness for 7 days. Number of oospores germinated in each mustard treatment was counted using a ‘Hemacytometer’ and BX41 light microscope (Olympus Corp., Tokyo, Japan). The experiment was performed as a CRD with four replications. The experiment as a whole was repeated four times.
Greenhouse trial.
Inhibitory effects of mustard plant exatracts on P. capsici crown infection in cucurbits were determined in the greenhouse trial. A sterilized mix of soil:peat:perlite (1:1:1) was used throughout the greenhouse trials. Seeds of ‘Eureka’ cucumber, ‘Magic Lantern’ pumpkin, and ‘Dickinson’ pumpkin were sown in 48-cell flats. Ten-day-old and 20-day-old cucurbit seedlings were transplanted into 15.2 cm-diameter plastic pots containing the above-mentioned sterilized mix. Similarly, mustard plants were grown in the greenhouse in 15.2 cm-diameter pots. Extracts of Thirty-five days old mustard plants were prepared in SDW. Three mustard treatments (FBL, Tilney, and a 50:50 mixture of FBL+Tilney) and a control (without mustard) treatment, were tested on P. capsici in the greenhouse trial. Zoospore suspension was prepared according to a method described by Babadoost and Islam (2002). Zoospore suspensions from the four isolates of P. capsici (PC-1, PC-2, PC-3, and PC-4) were equally mixed and number of zoospores in the suspension mixture was adjusted to 200,000 spores/ml. Cucurbit seedlings were inoculated by adding 10 ml zoospore suspension at the base of each cucurbit seedling. Immediately following the inoculation, 50 ml each of FBL, Tilney, FBL+Tilney and control (SDW) treatments were applied into the base of four different cucurbit seedlings. Seedlings were watered 3 times per day and development of P. capsici infection was assessed until 21 days after inoculating plants. The greenhouse trial was a factorial experiment in CRD with three replications. The greenhouse trial as a whole was repeated five times.
Field trials.
Four field trials, 2008-spring, 2008-fall, 2009-spring, and 2010-spring, were conducted in two commercial fields in Tazewell County near Pekin, Illinois. Both fields had Phytophthora blight on pumpkins in 2007. One field was used for 2008-spring, 2009-spring, and 2010-spring trials, and a second field was used for 2008-fall trial. The 2008-spring field trial was a split-split plot experiment performed in a randomized complete block design (RCBD) with three replications. The main plots included FBL, Tilney, and a 50:50 mixture of FBL+Tilney mustard and a control (no mustard) treatments. In each plot of FBL, Tilney, and FBL+Tilney, 806, 1,345, and 1,135 g of mustard seeds, respectively, was sown on 28 April using a seed broadcaster and covered with a steel bar harrow. Mustard plants were collected from the field immediately before the plants were incorporated into the soil. Three mustard plant samples (each from 1 sq m area) were collected on 13 June (45 days after sowing the seeds) from each plot and average fresh biomass (grams/m2) was determined. FBL mustard plants were at pre-bloom growth stage and Tilney mustard plants were at full-bloom, when mustard plants were collected. Mustard plants were incorporated into the top 10-cm layer of the soil using shallow disking and field cultivator on 13 June. Each main plot was divided into two sub-plots for two dates of sowing treatments of cucurbit seeds. Cucurbit seeds were sown on 13 June (the same day) and 17 June (after four days). Three sub-sub plots for three cucurbit cultivars ‘Eureka’ cucumber, ‘Magic Lantern’ pumpkin, and ‘Dickinson’ pumpkin, were included. The plots were harvested on 28 September.
In the 2008-fall trial, field was cultivated and prepared on 17 September. The exact same quantity of mustard seeds used in the main plots for spring-2008 trial, were sown on 22 September in 2008-fall trial. Three mustard plant samples (each from 1 m2 area) were collected on 23 October (30 days after sowing seeds) from each plot and average fresh biomass (grams/m2) was determined. Both FBL and Tilney were at pre-bloom growth stage, when mustard samples were collected. Mustard plants were left intact in the soil to overwinter. By early spring of 2009, all mustard plants were dead and almost completely disintegrated. The design of this field trial was similar to that of 2008-spring trial except that only one sowing date was included. This field trial was a split plot experiment in RCBD with three replications. The trial included four main plots (FBL, Tilney, FBL+Tilney, and control) and three sub plots (‘Eureka’ cucumber, ‘Magic Lantern’ Pumpkin, and ‘Dickinson’ pumpkin). Seeds of cucurbit crops were sown on 29 May, 2009. The plots were harvested on 3 September.
The 2009-spring and 2010-spring trials were conducted using the same design of 2008-fall trial. The design had four main plots (FBL, Tilney, FBL+Tilney, and control) and three sub plots (‘Eureka’ cucumber, ‘Magic Lantern’ Pumpkin, and ‘Dickinson’ pumpkin). In 2009-spring trial, mustard seeds were sown on 9 June and mustard plants were collected on 15 July (35 days after sowing mustard seeds). Three mustard plant samples (each from 1 m2 area) were collected from each plot and average fresh biomass (grams/m2) was determined. FBL plants were at pre-bloom growth stage and Tilney mustard plants were at full-bloom, when mustard samples were collected. Mustard plants were incorporated into the soil on 15 July using shallow disking and field cultivator. Seeds of cucurbit crops were planted on 16 July. The plots were harvested on 18 October. In 2010-spring trial, mustard seeds were sown on 26 April and mustard plants were collected on 1 July (65 days after sowing seeds). Three mustard plant samples (each from 1 m2 area) were collected from each plot and average fresh biomass (grams/ sq m) was determined. Both FBL and Tilney mustard plants were at full-bloom, when mustard plants were collected. Mustard plants were incorporated into the soil on 1 July using shallow disking and field cultivator. Seeds of cucurbit crops were planted on 2 July. The plots were harvested on 5 October.
From the mustard plants collected to determine biomass of each treatment, a subsample was used for quantification of glucosinolates. Mustard plants collected were washed to remove soil particles from roots. Mustard plant tissues were cut, frozen immediately in liquid nitrogen, and lyophilized. After lyophilizing, mustard tissues were ground using the coffee grinder (Mr. Coffee, IDS 55) and immediately transferred to -80ºC freezer. The ground mustard tissues were analyzed to quantify glucosinolates using high performance liquid chromatography (HPLC). Mustard samples were analyzed to quantify glucosinolates according to the procedure described by Kushad et al. (2004). The type and amount of glucosinolates, were estimated based on retention times and response factors developed for desulfoglucosinolates in the standardized and certified rapeseed reference material BCR 367 (Commission of the European Community Bureau of References, Brussels, Belgium).
There were a total of 72 plots in the 2008-spring trial and a total of 36 plots each in the 2008-fall, 2009-spring, and 2010-spring field trials. Twelve cucurbit plants were maintained in each plot (10.67 m× 9.14 m). Data were taken from these four field trials in biweekly intervals, starting from four weeks after planting cucurbit seeds. Infection by P. capsici and Fusarium spp. were visually assessed by observing seedling death, vine lesions, and fruit infection of cucurbit plants. In each plot, twenty vines and all the fruits were examined for the infection. Laboratory, greenhouse, and field data were analyzed using GLM procedure of SAS 9.2 (SAS Institute, Inc., Cary, NC). Fishers protected LSD was used to compare treatment means at P=0.05.
Research results and discussion:
Laboratory experiments.
Colony growth of P. capsici was significantly reduced, when the cultures were exposed to the extracts of FBL or FBL+Tilney, in comparison to the control treatment. Mean growth suppression of P. capsici isolates were 81.33, 8.05, and 65.88 % in FBL, Tilney, and FBL+Tilney mustard treatments, respectively compared to control treatment.
Exposing Fusarium cultures to the mustard extracts did not significantly affect the growth of the colonies. Mean colony radius of Fusarium isolates were 3.0389, 2.8333, 2.9278, and 3.0417 cm in FBL, Tilney, FBL+Tilney, and control treatments, respectively. Colony radius of F-2 isolate in Tilney treatment was reduced by 20% and colony radius of F-4 isolate in FBL+Tilney treatment was reduced by 11% in comparison with the control plates (Table 3.2). Isolate F-1 did not grow well in the culture plates.
Mean number of sporangia produced in four isolates of P. capsici per viewing field of the microscope were 67.96, 161.56, 73.83, and 163.51 in FBL, Tilney, FBL+Tilney, and control treatments, respectively. The FBL and FBL+Tilney treatments significantly reduced the sporangial production of PC-1 isolate compared to Tilney and control treatments. Also, FBL, Tilney, and FBL+Tilney treatments significantly reduced sporangial production in PC-3 isolate compared to the control treatment. No mustard treatments significantly affected the sporangial production in PC-2 and PC-4 isolates.
Mean percentages of zoospore germination were 75.11, 70.27, 81.07, and 67.40% in FBL, Tilney, FBL+Tilney, and control treatments, respectively. Zoospore germination of P. capsici isolates was not significantly affected by any of the mustard treatments.
Mean percentages of oospore germination were 40.61, 52.99, 43.33, and 57.10% in FBL, Tilney, FBL+Tilney, and control treatments, respectively. The FBL and FBL+Tilney treatments significantly reduced percentage of oospore germination compared to control and Tilney mustard treatments.
Greenhouse trial.
No crown infection was observed in ‘Eureka’ cucumber. All ‘Magic Lantern’ pumpkin plants exhibited crown infection. The rates of crown infection in ‘Dickinson’ pumpkin ranged from 25.23% (FBL+Tilney treatment) to 50.69% (control treatment). The FBL and FBL+Tilney treatments significantly reduced the P. capsici crown infection in ‘Dickinson’ pumpkin compared to control treatment.
Field trials.
Mean biomass of FBL, Tilney, FBL+Tilney treatments in four field trials were 2,016.66, 2042.50, and 1,874.17 g/ sq m, respectively. Mean biomass of 2008-spring, 2008-fall, 2009-spring, and 2010-spring trials were 2,965.56, 211.11, 1,975.56, and 2785.89 g/sq m respectively. In 2008-fall trial, all of the mustard treatments had lower biomass compared to other field trials. Glucosinolate content (µmol/sq m area) of mustard treatments in 2008-fall trial were the lowest and were significantly lower than 50% of the total number of treatments. The lowest glucosinolate content (31.19 µmol/sq m area) was in FBL+Tilney treatment in 2008-fall trial and the highest glucosinolate content (3,204.26 µmol/sq m area) was in FBL+Tilney treatment in 2009-spring trial. Average glucosinolate content in the fresh tissues of FBL, Tilney, and FBL+Tilney treatments in four field trials were 1,240.99, 916.75, and 1,417.63 µ mol/sq m, respectively. Average glucosinolate content of 2008-spring, 2008-fall, 2009-spring, and 2010-spring trials were 1,636.80, 286.59, 1,482.09, and 1,552.73 µ mol/sq m, respectively.
Vine infection was not observed on “Eureka’ cucumber. The rates of infected vines of ‘Magic Lantern’ and ‘Dickinson’ pumpkin combined were 19.11, 25.91, 22.96, and 19.33% in FBL, Tilney, FBL+Tilney, and control treatments, respectively. In none of the field trials, mustard treatments significantly reduced the rates of vine infection by P. capsici compared to control treatment. Overall mean percentages of vine infection in ‘Magic Lantern’ pumpkin and ‘Dickinson’ pumpkin were 27.95 and 18.95 %, respectively. No vine infection by Fusarium was detected in any of the experimental crops.
The rates of fruit infection by P. capsici were 29.48, 23.20, 21.71, and 26.32% in FBL, Tilney, FBL+Tilney, and control treatments, respectively. Percentages of fruit infection in 2009-spring treatment were the lowest among the four field trials. Overall rate of fruit infection in ‘Eureka’ cucumber, ‘Magic Lantern’ pumpkin, and ‘Dickinson’ pumpkin were 25.74, 23.77, and 20.32%, respectively, which were not significantly different from each other. No Fusarium infection on any fruit was observed.
DISCUSSION
In the laboratory study, extracts of B. juncea (FBL) significantly suppressed colony growth of P. capsici isolates compared to that of S. alba (Tilney). Larkin and Griffin (2007) also reported that B. juncea completely inhibited the colony growth of Phytophthora erythroseptica while S. alba inhibited nearly 26% growth of P. erythroseptica. These findings indicate that glucosinolates present FBL can suppress colony growth of Phytophthora spp. In the laboratory experiment, growth of P. capsici colony was suppressed as long as the culture plates were exposed to mustard extracts. When the culture plates were separated from the plates containing mustard extract, the colonies started growing. Fan et al. (2008) reported that extracts of B. oleracea var. caulorapa (Kohlrabi) suppressed colony growth of P. capsici, but the suppression was not persistent. The result of our study and result of Fan et al. (2008) indicated that the effect of glucosinolates on growth of P. capsici is temporary and lasts as long as the fumigation effect is present. Further studies are needed to determine the suppressive effects of individual glucosinolates in Brassica species on colony growth of P. capsici. Also, other mustard cultivars should be utilized to test their inhibitory effect on colony growth of P. capsici.
The results of our study showed that production of P. capsici sporangia was significantly suppressed in two isolates by FBL extract and in one isolate by Tilney extract. These findings indicate that there are interactions between glucosinolates and P. capsici isolates. Further studies are needed to determine the effectiveness of each glucosinolate on different isolates of P. capsici. Tilney treatment did not significantly affect germination of P. capsici oospores. The FBL treatment reduced germination of P. capsici oospores by only 29% in comparison with control. Yet, 40.61% of P. capsici oospores germinated in FBL treatment. Also, the result of our study showed that germination of P. capsici zoospores was not significantly affected by any of the mustard treatment. Thus, the use of mustard as a cover crop for management of P. capsici in the field should aim at suppressing hyphal growth and sporangial production rather than oospore and zoospore germination.
None of the cucumber seedlings in the greenhouse study and none of the cucumber vine in the field were infected by P. capsici. This finding did not agree with the report by Tian and Babadoost (2004) that all of the cucumber seedlings were infected by P. capsici. The cucumber cultivar used in the study of Tian and Babadoost was different from the cultivar (‘Eureka’) that we used in our study. ‘Eureka’ cucumber is likely to be carrying resistant gene(s) against vine infection by P. capsici, which is not resistant to fruit infection by P. capsici. The FBL treatment reduced seedling infection of ‘Dickinson’ pumpkin by 38%, but FBL treatment did not significantly affect infection of ‘Magic Lantern’ pumpkin seedlings in the greenhouse trial.
None of the mustard treatments significantly reduced the vine infection of either ‘Magic Lantern’ pumpkin or ‘Dickinson’ pumpkin in the field trials. So, using either FBL or Tilney, or a combination of these mustard cultivars as cover crop is not an effective method for management of vine infection of pumpkins. In addition, fruit infection of either “Eureka’ cucumber, ‘Magic Lantern’ pumpkin or ‘Dickinson’ pumpkin was not significantly affected by mustard treatments in the field trials. Thus, effective control of P. capsici using mustard as a cover crop alone appears to be unlikely. However, utilizing mustard as a cover crop in an integrated approach (for example, in combination with fungicide application and crop rotation) for managing P. capsici in cucurbits may be a viable approach.
The lowest biomass was in the 2008-fall trial. Mustard plants in 2008-fall trial were not growing as fast as the mustard plants of same age in other field trials. Field temperature might be the main reason for the slow growth of plants in 2008-fall trial. For the growth and development of B. juncea and S. alba, the plants need certain threshold level of heat units represented as growing degree days (GDD) (Miller et al., 2001). The accumulated GDD (calculated at base 50°F) were 573, 340, 820, and 1182 in 2008-spring, 2008-fall, 2009-spring, and 2010-spring trials, respectively. For the 2008-fall grown mustards, the low GDD might have reduced the rapid growth of the plants in the field. In the spring field trials, morphological differences and growth characteristics of FBL and Tilney might have contributed to the difference in the biomass in different trials. Additional studies needed to be done to determine the biomass of FBL and Tilney at different growth stages. Biomass influenced glucosinolate content (µmol/m2) in the mustard plants incorporated into soil. Higher biomass of 2008-spring, 2009-spring, and 2010-spring trials compared to 2008-fall trial contributed to higher glucosinolate content (µmol/m2) in the mustard plants incorporated in those trials compared to 2008-fall trial. Considering the glucosinolate content (µmol/m2), the best time to incorporate FBL mustard cultivar was between 45-65 days. Also, the best time to incorporate Tilney mustard cultivar was between 35-45 days.
Sowing cucurbit seeds on the same day of mustard incorporation into the soil did not reduce their germination compared to sowing cucurbit seeds four days after mustard incorporation. Thus, cucurbit seeds can be sown any time after mustard incorporation in to the soil without significant effect on either seed germination or plant vigor. Fall field trial was not repeated during 2009 and 2010 seasons. For fall trial, fields were only available from the late September (after harvesting crops). Plants are usually killed by frost in early November. As a result of these factors, sufficient biomass is not produced during fall seasons. Thus, fall trial of mustards was discontinued after 2008 season.
The overall conclusion of the laboratory, greenhouse, and field studies during the three- year period showed that using mustard cultivar alone for management of P. capsici in cucurbits is not adequate. However, we tested only two cultivars of mustard. Testing of other mustard cultivars may have different results on the effectiveness of mustards for control of P. capsici in cucurbits.
Project outcomes:
The results of laboratory and greenhouse trials showed a good potential for using short-cycle mustard cover crops for managing Phytophthora blight in cucurbits. But, the results from the field studies were not consistent. With the available results, the recommendation would be using mustard cover crop in an integration approach with cropping rotation (with non-host crops) and fungicide applications for management of Phytophthora blight in cucurbits.
Economic Analysis
Using cover crops in cucurbit production is a common practice. Short-cycle mustards can be grown to maximum capacity in about 40 days. Thus, using a short-cycle mustard in early spring as a cover crop, prior to planting cucurbits, is feasible. The cost for purchasing seed, sowing seed, and incorporating plants into the soil are very affordable. In deed, some growers have been using mustards as cover crops. However, using any cover crop, including short-cycle mustards, in the spring is not feasible for processing pumpkins, as processing pumpkins are planted as soon as weather conditions in spring permitsFarmer Adoption
A few squash and jack-o-lantern pumpkin growers have tried short-cycle mustards as cover crop in their farms in Illinois. However, since the field results have not been consistent, wide adoption of mustards as cover crops has not been established yet.Recommendations:
Areas needing additional study
There is a good potential of using short-cycle mustards as cover crops for management of soil-borne fungal pathogens, including Phytophthora capsici and Fusarium spp. More research are needed to determine effectiveness of mustard cultivars for this purpose, including: 1) testing all potential mustards lines/cultivars and selecting the most effective ones in suppressing soil-born pathogens; 2) screening or breeding for shortest cycle mustards (< 30-day mustard) as cover crop; 3) investigating effectiveness of fall-grown mustard vs effectiveness of spring-grown mustards; 4) establishing regional research for determining effectiveness of mustards as cover crops for managing soil-born pathogens; and 5) determining the value of mustard as a cover crop in integration with other practices (e.g., cropping rotations and chemical use) for managing soil-born pathogens.{kind=link}
{kind=link}