Final report for GNE24-316
Project Information
Background and Project Goals
In agriculture, the mutualisms that ants and other predatory insects share with plants that produce extrafloral nectar (EFN) is understudied1. Extrafloral nectaries (EFNs) are structures that some plant groups produce that secrete nectar for the purpose of attracting omnivorous insects in exchange for protection from herbivores2. Ants are the most important insect group involved in this interaction, but other predatory insects visit EFNs and provide protection2–4. Conservation biocontrol is the concept of promoting habitat on farms that is attractive and hospitable to native beneficial insects, which provide essential services like pollination and pest removal. I aim to contribute to the knowledge of using EFN-producing plants as a means for conservation biocontrol in organic New England agroecosystems. Here, I outline a project designed to determine the effectiveness of utilizing EFN-producing companion plants for the benefit of cucumber production in an organic farm setting.
Research and Education
I used partridge pea (Chamaecrista fasciculata), an EFN-producing wildflower, as a companion plant to grow alongside cucumber. During the summer of 2025, my colleagues and I collected insect community, cucumber leaf damage, and harvest data throughout the growing season. These data were compared across replicate blocks of the following three treatments: partridge pea, mulched ground (partridge pea-free control), and weedy fallow. My outreach component consisted of two parts: 1) utilizing and dispersing informational materials provided by the Xerces Society while hosting informational booths at local farmers markets and 2) insect collection and education events with a summer youth program (Recreation Worcester) in various urban parks in Worcester, MA.
Main results and conclusions
Overall, the inclusion of partridge pea did not significantly increase the yield of cucumber. However, partridge pea treatments were on par in terms of productivity compared to control cucumber plots, with the weedy treatment being less productive than both partridge pea and control treatments. Weedy fallow likely competes with cucumber for nutrients and provides few synergistic effects that outweigh the detriments. Whereas partridge pea was extremely productive in floral and extrafloral rewards that likely allowed for higher abundances generalist predators like wasps and ladybeetles. There were also fewer aphids collected near cucumber that were growing next to weeds and partridge pea compared to controls Additionally, our cost-benefit analysis showed no significant reduction in potential profit gained when including costs associated with including partridge pea. These results suggest that partridge pea may be a viable companion plant in organic cropping systems. Information gained from studies like mine builds on our understanding of the plant-protecting activities that abundant omnivorous insects like ants, beetles, and wasps provide, which is an incredibly useful tool for farmers dedicated to sustainably growing food.
Impacts, challenges, and unexpected outcomes
There were some unexpected results from this study that conflicted with my initial predictions. Broadly, I expected cucumbers growing next to partridge pea to be more productive than control, but this study was limited to one summer at two separate farms. Any net gains in cucumber productivity are likely to be seen over a larger time scale, as it may take multiple growing seasons for beneficial arthropod and nitrogen-fixing bacteria to establish in an area. Ant communities and pollinators did not significantly vary among treatments, which I expected them to. This may also be due to the scope of the project being limited to one year and two sites. These findings may also be accurate, and these specific insect groups (while very abundant and diverse on the partridge pea themselves) may not be as willing to forage on the nearby cucumber.
- Examine the effects of EFN-producing companion plants on insect communities associated with organic cucumber plantings in New England.
- Determine how EFN-producing companion plants influence the presence of pests, crop damage, and yield of cucumber plantings.
- Determine whether companion plants influence crops with and without their own EFNs differently.
The purpose of this project is to expand our current understanding on the utilization of plants that produce extrafloral nectaries (EFNs) as a tool for conservation biocontrol in organic vegetable systems. Extrafloral nectar evolved to reward omnivores like ants with sugars in exchange for defense against herbivory2,5. While ants are the primary mutualist, other beneficial predators known to feed on EFNs and reduce herbivory include parasitoids, beetles, lacewings, and even spiders2–4. Past studies have indicated the use of EFNs as a promising method of recruiting beneficial predatory insect abundance in agroecosystems, but their efficacy is highly dependent on a farm’s management practices, crops, and the surrounding ecosystem1. Therefore, I explored methods of using native EFN-bearing plants to harness the behavior of local ants and other predaceous omnivores to advance pest control for organic growers in New England.
Partridge pea (Chamaecrista fasciculata) is an EFN-producing legume native to the Eastern US that is highly attractive to predatory and pollinating insects. The use of this plant in agroecosystems is understudied, but they have great potential for enhancing the beneficial arthropod community in an area6. Here, I focused on cucumber (a non-EFN cucurbit), which is popular among organic farmers in New England. I used partridge pea as a companion plant treatment alongside cucumber and assessed its effect on insect community dynamics and cucumber yield. The combination of plants in this system are good study candidates for this project. Cucumber is host to two major pests of economic concern to New England farmers: striped cucumber beetles (Acalymma vittatum) and spotted cucumber beetles (Diabrotica undecimpunctata howardi), which vector bacterial wilt that can cause up to 20% yield losses if left unmanaged by farmers7. Partridge pea intercrops may attract parasitoids and generalist predator insects (like ants) that could assist in controlling the populations of these beetles and their subsequent damage - which is what I aimed to assess.
There is an increasing societal demand for organic produce and less reliance on chemicals in food production8,9. In addition to consumer concerns, the use of broad-spectrum insecticides also harms non-target beneficial organisms like pollinators and predaceous arthropods10. Utilizing the prey-regulating ability of the predatory insects could help us mitigate crop damage, while reducing our reliance on these chemicals. Releasing lab-reared biocontrol organisms is common but is expensive and releasing species outside their native ranges can negatively non-target arthropods, especially endangered local species11,12. I would like to expand on this concept and learn more about the possibilities of enhancing local beneficial insect populations via the attraction of EFN-producing plants native to New England.
This proposal is part of my dissertation at Clark University, where I study the roles of ants in agroecosystems. The experiment outlined here is part of my second chapter, where I aim to run field experiments using cultivation methods common among organic farmers in Massachusetts. For the summer of 2024, I conducted my first set of trials for this chapter by utilizing partridge pea as a companion plant alongside an EFN-producing cucurbit vegetable: zucchini. The purpose of this study was to determine if the inclusion of an EFN companion plant will attract and support nearby ant/beneficial arthropods, which would then spillover and provide their predatory services to the zucchini crops. Due to limitations in budget and labor, this study took place at a single site. The experiments outlined in this proposal took place during the summer of 2025, and were not simply an extension of my 2024 dataset, rather they focused on a new crop, included a weedy fallow treatment, and took place in larger plots at two separate sites.
I aim to provide New England cucurbit growers with new information on this understudied and potentially effective chemical-free alternative to pest control, enhancing our current understanding of the mutualism between plants that produce EFNs and the organisms they reward in exchange for defense. To date, there has not been an extensive study on the employment of ants as biological control in cucurbits via intercropped EFN recruitment1. As our population expands, food security issues will become more common, especially in other regions of the world where factors like global change and poor soil quality are likely to hit the hardest13,14. Developing new conservation biocontrol practices will reduce the need for costly pesticides and horticultural practices that create high barriers of entry that may dissuade people without the necessary capital for those tools from starting their own farms. Any work that supports local farmers improves the surrounding community, helps eliminate food deserts in poor areas, and is a step toward phasing out our dependence on large food distribution corporations that we currently rely heavily on for produce.
Cooperators
- (Researcher)
- (Researcher)
- (Researcher)
- (Researcher)
- (Researcher)
- (Educator)
- (Educator)
Research
Study area
This field experiment took place in Massachusetts on two sites. I utilized 0.20 acres at both sites for this study (Fig. 1), which allowed for ample space between treatment blocks and a mowed perimeter. For the summer 2024 season, I worked with a land manager in Spencer, MA (Ronald Audette in the cooperators listed here) and I continued to utilize this space in 2025. A board member of the Grafton Land Trust (Paul Grady in our cooperator section) allowed me to use part of their pasture that they manage for the same purposes. Farms in Central MA share common pests and beneficial species, yet each farm can vary in their insect community composition and management strategy (personal observations 2022-2025).
Cucumber is the focal crop of this study. I constructed 24 equally-sized blocks at each site acting as individual replicates (Fig. 2). Each block was 3 x 5m, allowing for multiple rows of crops and companion plants with space for 3m mowed buffer areas between blocks to reduce the interactions among blocks of different treatments. One block consisted of one 5m long central crop row and an equal-length treatment zone on one side (Fig. 3). In a randomized block design, these blocks received one of three possible planting treatments: 1) control treatment only surrounded by weedy fallow, 2) bare mulched ground, 3) cucumber with partridge pea (Fig. 4). There were four replicates of each treatment at each site, for a total of eight replicates of each treatment. For all analyses, the sample size equals the number of experimental units being sampled (48). I treated all rows with the same watering regimes. The experiment will end three weeks after peak cucumber harvest.
Treatment justifications
Cucumber was chosen as the focal crop because it is an economically important vegetable for organic farmers in our area. The weedy fallow control treatment is necessary in order to use as a reference point for unmanaged habitat that acts as refuge for local arthropods, which can be compared to to a planting intentionally selected for its properties as an insectary crop15,16. The bare ground treatment allows me to compare our treatments to conventional cucurbit production (e.g., mulched/weedless). Partridge pea was chosen as the companion plant treatment due to its great potential as an understudied attractor of beneficial arthropods6. Additionally, partridge pea is a legume which can be utilized for soil nitrogen fixation, and its flowers are highly attractive to native pollinators (personal observations 2022-2024) which may prove an additional benefit of increased fruit set for cucumber.
Objective 1
I aimed to determine the effectiveness of EFNs as attractors of ants (Fig. 5), other predators, as well as pollinators (Fig. 6). I addressed this objective by collecting herbivore and ants/beneficial arthropods to determine their density and diversity among treatments. Insect presence (both pests and predators) on cucumber plants during morning visual observations was minuscule this year, so I instead decided to focus on traps for estimating insect populations rather than in-person observations and vacuum sampling. Insects were collected throughout the growing season using various methods to obtain a representative sample of the insect community from each block. These methods included pitfall traps, sticky cards, and baited traps (tuna/honey)(Fig. 7). Collecting and preserving insect specimens this way is beneficial as I can identify individuals much further down taxonomically than with visual observations. I recorded environmental variables known to influence insect communities during each sampling date, including: soil temperature, soil moisture, soil pH, soil compaction, air temperature and relative humidity (iButtons), overhead canopy cover, precipitation, and plant volume. These environmental factors are necessary to record because each may vary by block and site, and accounting for them in analyses is crucial. During leaf damage and disease measurements, I recorded the presence and absence of wingless aphids on the underside of each leaf, and included it as a factor in subsequent analyses.
I then used data generated from these traps to calculate the richness and/or abundance of ants, syrphids, lady beetles, ground beetles, generalist wasps, parasitoid wasps, aphids, striped cucumber beetles, and bees. Ground-dwelling beetles trapped in pitfalls were identified to family and quantified. Carabids were the most numerous beetle family, and are also considered important generalist predators on farms. Thus, their abundance was analyzed further. I used linear mixed effects models to examine how these specimen groups are impacted by treatment groups and environmental variables. I also conducted Non-metric Multi-Dimensional Scaling (NMDS) analyses on the presence/absence data gathered from the different trapping methods to compare insect community composition between the planting treatments and sites. I predicted that partridge pea will increase abundances of generalist omnivores like ants and beetles on focal crops, as well as parasitoid wasps/flies whose adults feed on EFNs, and that crop damage will be lowest and yield highest in this treatment8. I also predicted that weedy fallow treatments will provide plant structure and serve as a refuge for predatory insects, but will not be as attractive as the partridge pea blocks - and therefore this treatment will have lower beneficial insect diversity than the partridge pea treatment.
Predictions regarding aphids
Ants are known to tend aphids in exchange for honeydew, which increases aphid numbers on plants. However, I did not predict that enhancing ant populations via EFN companion plants would be an issue regarding aphids in my study system. Aphids, while present on cucumber and zucchini, are not the main cucurbit pest of concern in New England. Stripec cucumber beetle (Acalymma vittatum) is the most damaging pest of cucurbits growing in our region17, and I have witnessed the effects of this pest firsthand when conducting my first two seasons of samples across organic farms in Massachusetts. Net economic impacts of ant-aphid mutualisms are highly context dependent, but studies in other systems have shown positive effects of honeydew-farming ants that drive away other arthropod pest groups from plants harboring aphid colonies18. Lastly, EFNs and flowers of my companion plants are highly attractive to local wasps that parasitize aphids, as well as voracious aphid-eaters like hoverflies, ladybeetles, and lacewings6,19.
Objective 2
I sought to determine if EFN-bearing companion plant presence results in less crop herbivory and higher yield compared to controls. Here I compared crop yield and leaf damage/disease among the treatments described above. I obtained biomass by weighing produce as it was harvested. I quantified leaf damage by scoring a subsection of each plants’ leaves by level of damage after practice via the online tool: https://zaxherbivorytrainer.com. Disease was scored as either being present or absent on leaves while simultaneously examining for damage. The inclusion of these categories is important, as different types of damage indicate different pests/disease. Schifani et al., 2020 suggested that the presence of ants on experimental pear trees lowered necrotic damage caused by a fungus, via antimicrobial secretions from visiting ants20. Therefore, recording the percentage of plants infected with disease will provide useful information. Lastly, I will use yield data to conduct a cost-benefit analysis to determine if the materials required to install/maintain these companion plantings are worth it to a farmer. Our produce was donated to local food fridges (https://www.woofridge.org/), but I calculated hypothetical profits by using the average price/lb of cucumber provided by the USDA (https://www.ers.usda.gov/data-products/fruit-and-vegetable-prices), and then subtracted the estimated cost of planting partridge pea (using cost of seeds, legume inoculant, and labor to seed these rows).
I hypothesized that the recruitment of predators to EFN companion plants will influence the marketable crop yield and tissue damage in a system. I predicted that when compared to control blocks, there will be an increase in marketable crop yield and lower host plant tissue damage from herbivores when partridge pea are present. This is because partridge pea should attract higher numbers of generalist predators (like ants) that I hypothesized are important in removing the most damaging pests in cucurbits (e.g., A. vittatum, D. howardi)21,22.For the cost-benefit analysis, I predicted that the yield gains from partridge pea will outweigh the overall cost of materials (e.g., seeds) and labor to include these plantings when compared to the management that goes into maintaining control blocks.
Objective 3
Zucchini is a cucurbit with similar growing requirements and seasonality to cucumber. One important difference is that zucchini bears EFNs on the underside of their leaves, while cucumber does not. For my last objective I determined if the existence of EFNs on cucurbits being produced has an impact on the ants (the most numerous EFN visitor in our studies), important pests, and overall yield of these closely related cucurbits when grown alongside partridge pea. I accomplished this by comparing the ant community composition, ant species richness, and the abundance of striped cucumber beetles (the most economically-important cucurbit pest in New England) between cucumber grown in 2025 and zucchini grown in 2024. Due to limitations during the summer of 2024 prior to my application for the SARE grant, I was only able to grow zucchini at one site and with two treatments (i.e., no weedy treatment). Therefore for consistency, only cucumber data from the same site in 2025 (Demer’s Hill in Spencer, MA) was included in these analyses comparing the two cucurbits, and the weedy treatment was excluded. I grew zucchini and collected data under similar experimental methods as outlined in Objective 1.
I hypothesized that ants and striped cucumber beetles would vary between zucchini and cucumber, and among combinations of their companion plant treatments. I predicted that carbohydrate-seeking predatory arthropods that utilize EFNs (e.g., ants) will be more apt to spill over to and forage on zucchini versus cucumber. The presence of EFN on zucchini will cause ants to spend more time on this crop versus its non-EFN counterpart (i.e., cucumber). For zucchini specifically, I predicted that more ant species will forage on this crop’s EFNs (and therefore more will be collected from these plants) in “PP only” versus other treatments, as this treatment will attract and host the most ants. Predictions for zucchini yield were similar to those of cucumber above in Objective 2, with higher yields from the partridge pea treatment than the controls.
Sharing of data and results
I will publish the results from this study in relevant scientific journals as part of my dissertation, as well as present to (e.g., local NOFA symposiums) and share with (e.g., data reports) farmers, landowners, and groups involved in the outreach presented in this proposal.
Statistical analyses
Various statistical analyses were used to answer each specific question in Rstudio version 4.5.223. All data were first tested for violations of the assumptions of their appropriate analyses. Unless noted otherwise in the results section, most data were analyzed with linear mixed-effects models (LME) (lmer function in the lme4 package) when possible, model residuals were tested for normality by viewing quantile-quantile plots24. Poisson generalized linear mixed-effects models (GLMM) (glmer function in the lme4 package) were used if LME residuals were still non-normal after transformation, as some datasets were heavily zero-inflated. GLMMs were tested for assumptions of overdispersion using the DHARMa package in R25. Environmental factors were chosen as main effects (alongside experimental treatment) by removing those that were highly correlated in pairwise tests after constructing spearman correlation matrices (cor function in the stats package)23. Random effects (experimental artifacts like location, sampling date, block number, etc.) were included in the full models and only removed if their effect sizes were zero or near zero to avoid model singularity (ranef function in the lme4 package). Specific model formulas for each analysis are noted in the output tables provided. When significant effects of a categorical variable were detected in a model, the emmeans function from the emmeans package was utilized to conduct post-hoc tests26. Where necessary, omnibus tests are displayed in the results section to show overall statistical differences between categorical fixed effects with more than 3 levels when found to be significant in LME and GLMM analyses.
I also used Non-metric Multi-Dimensional Scaling (NMDS) (metaMDS function from the vegan package) analyses to compare the ant and insect community compositions between companion planting treatments (Objective 1), and ant species across crop types (Objective 3)27. These analyses allow me to visually determine if the composition of species (i.e., the presence and absence of certain groups) differs from one treatment to another by calculating Jaccard dissimilarity values and plotting them on two axes. The significance of these differences were determined using Permanovas (adonis2 function from the vegan package).
Fig. 1: Plots for our 2025 study.

Fig. 2: Plot overview
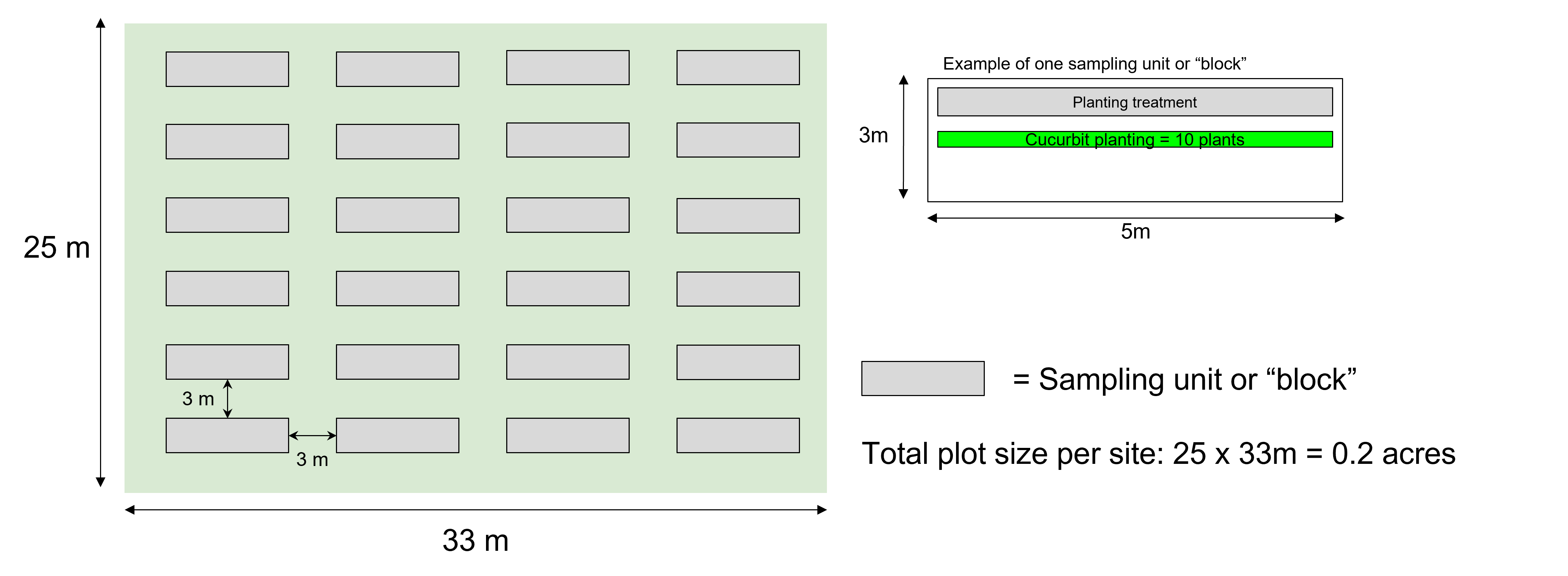
Fig. 3: Plot closeup
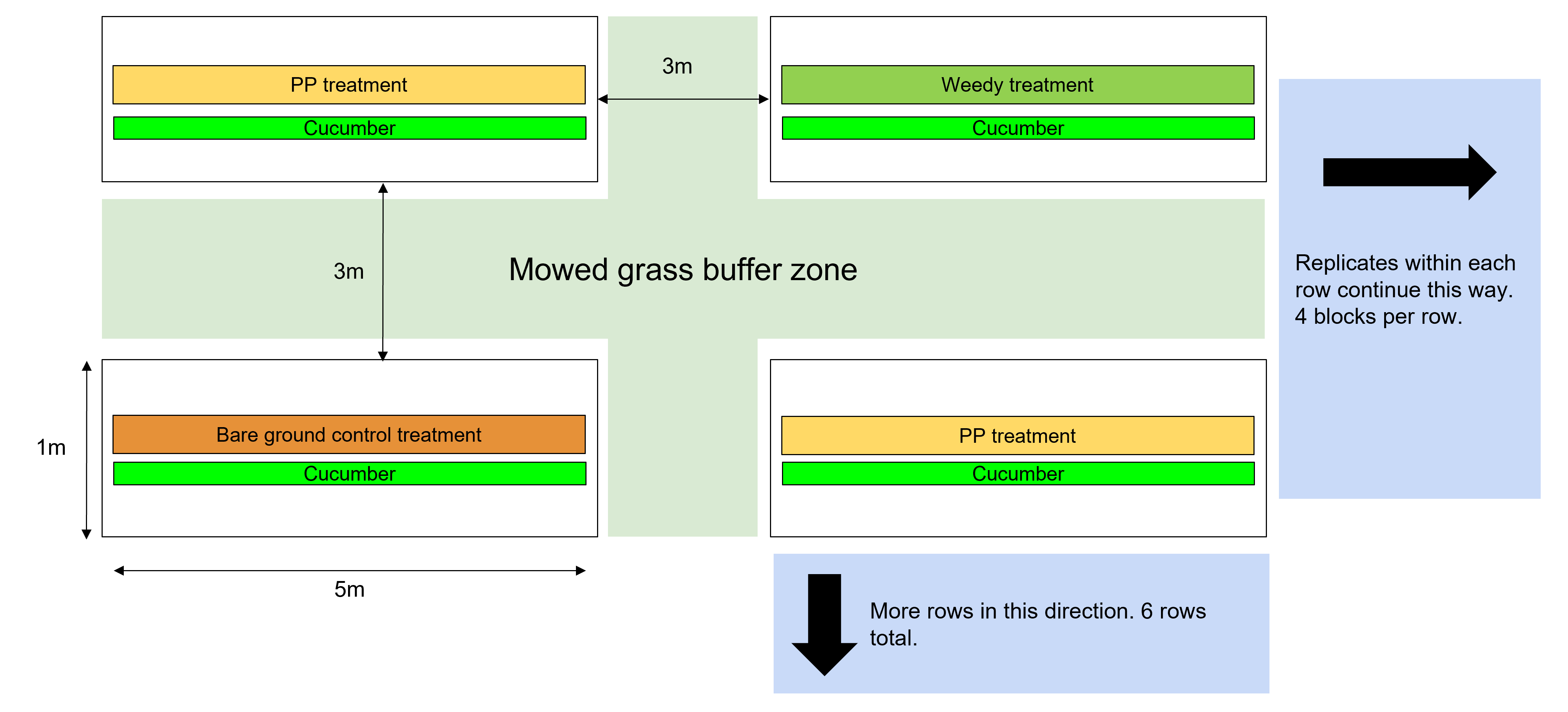
Fig. 4: Examples of each planting treatment for cucumber

Fig. 5: An ant foraging on one of our partridge pea EFNs

Photo credit: Steven King
Fig. 6: A bumblebee foraging on one of our partridge pea flowers

Photo credit: Steven King.
Fig 7. Various insect trapping methods used in this study

I generated many tables and figures for these analyses. Below, I have attached them in a PDF document. Any Table or Figure labeled with "S" in the results section below refers to the materials in this document.
Nelsen SARE 2026 supporting info
Objective 1
Insect community dynamics among the treatments
Ants
Tuna, honey, and pitfall data were combined when analyzing ant species richness. Richness was the only measurement used for ants, as their social recruitment behavior makes it impossible to examine abundance and diversity from traps (i.e., a single worker’s trail will lead others from that colony into the trap, thus inflating the count of that species). There were significant differences in the number of ant species across the three treatments from the main model (Anova: F(2, 182.9) = 3.317, p = 0.039). Specifically, control plots had significantly more ant species than partridge pea and weeds (Table S1). However, the adjusted p-values were no longer significant after conducting a post-hoc analysis using estimated marginal means when comparing control to partridge pea treatments (p = 0.084) and control to weedy treatments (p = 0.064) (Table S2). These near-significant differences are possibly due to differences resources provided by the weedy and partridge pea treatments that favor few, but dominant, ant species. There was also a significant effect of the presence of aphids on leaves, with more ant species being collected in plots that had aphids present (Anova: F(1, 183.30) = 4.579, p = 0.034). This result is not too surprising, as the presence of aphid honeydew likely allows for better resource partitioning among ant species in a plot. There were no significant correlations between ant species richness and soil compaction (Anova: F(1, 183.38) = 1.424, p = 0.235) or mean air temperature (Anova: F(1, 182.88) = 0.1094, p = 0.741).
After conducting an NMDS and Permanova to determine differences in ant community structure, I found significant differences between treatments (Permanova: F(304, 306) = 2.8876, R2 = 0.019, p = 0.0026) (Table S3). Specifically, control plots significantly varied from weedy (p = 0.049) and partridge pea (p = 0.010) (Table S4). However, viewing the ordination plot shows that the polygons of these two treatments mostly overlap, and the differences seen are likely driven by a few uncommon ant species that were collected from one of these treatments, but not the other (Fig. S1).
Ground beetle richness and Carabid abundance
Beetle family richness varied by treatment (Anova: F(2, 178.91) = 4.7975, p = 0.009), and was significantly higher in weedy versus control plots (p = 0.008) (Tables S5,S6)(Fig. S2). Beetle family richness was not correlated with soil compaction (Anova: F(1, 165.70) = 0.161, p = 0.688822) but had a significant negative correlation with soil pH F(1,106.67) = 5.515, p = 0.020697, partial r-squared = 0.032). Carabid beetle abundance was significantly influenced by treatment (Anova: F(2, 178.76) = 5.5413, p = 0.004625), but not soil compaction (Anova: F(1,156.84) = 0.750, p = 0.38), or pH (Anova: F(1, 172.59) = 3.443, p = 0.065). There was a significant influence of treatment on Carabid beetle abundance in pitfalls (Anova: F(2, 178.76) = 5.541, p = 0.005), but not soil compaction (Anova: F(1, 156.84) = 0.750, p = 0.3878) or soil pH (Anova: F(1, 172.59) = 3.4431, p = 0.065). Specifically, there were more Carabids in weedy plots than controls (p = 0.005)(Tables S7 and S8)(Fig. S3). The significant influence of weeds on beetle richness and Carabid abundance was anticipated, as more refuge and higher plant diversity are likely to lead to more niche diversification of generalists like predatory beetles.
Sticky card insects
Predators (parasitoid wasps, syrphid flies, ladybeetles, rove beetles, vespid wasps), pollinating bees, and pests (striped cucumber beetles and aphids) caught on sticky cards were counted and analyzed. There were no significant effects of cucumber treatment (Anova: F(2, 179.333) = 2.348, p = 0.0985) or relative humidity (Anova: F(1, 6.476) = 5.387, p = 0.056) on the number of parasitoid wasps found on sticky cards. However, temperature was positively correlated with parasitoid wasp abundance (Anova: F(1, 41.952) = 4.629, p = 0.0372, partial r-squared = 0.038) (Table S9). This result was not anticipated, as I expected parasitoid wasps to be much more attracted to the EFNs provided by partridge peas. It is possible that these wasps were attracted, but there could have been less movement between the partridge peas and the crops they were planted next to. The sticky cards were low to the ground in between cucumber plants, and partridge pea EFNs were higher at about 1-3ft at the end of the growing season, and it is possible that the wasps were too far from the cucumber to forage on them.
There was a significant influence of treatment (Anova: F(2, 181.803) = 4.6754, p = 0.011), a positive correlation with temperature (Anova: F(1, 167.588) = 49.562, p < 0.0001, partial r-squared = 0.217), and a negative correlation with humidity (Anova: F(1, 94.407) = 7.733, p =0.007, partial r-squared = 0.112) on aphid abundance from sticky cards. Specifically, there were significantly more aphids trapped in cucumber control plots than both those containing partridge peas (p = 0.014) and weeds (p = 0.044)(Table S10 and S11)(Fig. S4). These results mostly align with my expectations. Control plots likely allow for easier detection of host plants (as there are no other plants around to screen the crops. Also, the significant increases of generalist predators like lady beetles and vespid wasps in the weedy and partridge pea treatments suggest that predation from these groups were keeping aphid numbers down on the cucumber in those treatments. The negative correlation between aphid abundance and humidity was unexpected, as I assumed insect activity would be more likely to be influenced positively to moisture. It is also worth noting though, that the partial r-squared value was somewhat low at 0.112
Striped cucumber beetles (A. vitattum) did not differ by treatment (Chi-squared = 4.1216, DF = 2, p = 0.127) or humidity (Chi-squared = 0.002 DF = 1, p = 0.969), but mean temperature was positively correlated with their abundance (Chi-squared = 12.613, DF = 1, p = 0.0004, trigamma partial r-squared = 0.07495742)(Table S12). I expected to see less striped cucumber beetles in the partridge pea treatments overall due to predation via omnivores. However, it is possible that their numbers were too low this season to detect any real differences that may have been caused by treatment effects (e.g., the striped cucumber beetle abundance data was heavily zero-inflated). Additionally, it is possible that the striped cucumber beetles were attracted to the refuge provided by partridge pea or weeds, counteracting any effects of their reduction via increased predation.
All pollinating bee groups (e.g., honeybees, bumblebees, sweat bees) were not numerous enough to be analyzed separately, and were instead grouped into a single category. Bee abundance was not influenced by cucumber treatment (Anova: F(2, 181.201) = 1.352, p = 0.261), temperature (Anova: F(1, 25.100) = 0.179, p = 0.676), or humidity (Anova: F(1, 3.239) = 6.451, p =0.079)(Table S13). I expected there to be more pollinating bees on sticky cards in cucumber growing next to partridge pea. One possibility is that because partridge pea flowers only produce pollen (no nectar) that these bees were less interested in foraging on the cucumber of this treatment. Generally, I saw many honey and bumblebees foraging on partridge pea flowers while sampling, so it is possible that they simply preferred the taller partridge pea flowers versus the cucumber flowers lower to the ground (and closer to the sticky cards). Another reason is that larger bees like bumblebees and honeybees are too strong to be effectively trapped on a sticky card, especially after rain when the cards were wet. One of the undergraduate students that assisted in sampling this year received a fellowship from our university to conduct a separate project of their own in our cucumber fields. For this side project, they placed bee bowls (multicolored bowls filled with soapy water) alongside our pitfalls. Bee bowls are much more effieicent at collecting large flying pollinators. They are still working on identifying these specimens and will present the results at an upcoming Clark poster session. Aquiring this data will help us understand more about how cucumber pollinators are influenced by the inclusion of partridge pea companion plantings.
Syrphid fly adult abundance did not significantly vary by treatment (Anova: F(2, 170.826) =0.623, p = 0.538), was positively correlated with temperature (Anova: F(1, 156.922) = 4.509 , p = 0.0353, partial r-squared = 0.020), but not correlated with humidity (Anova: F(1, 81.364) = 1.440, p =0.234)(Table S14). This was another result that I did not expect, as syrphid fly adults are known foragers of flowers (both nectar and pollen). However, like the results seen with bees, it is possible that since partridge pea flowers do not produce nectar, those plots are less enticing. Another explanation is that since syrphid adults will lay eggs on plants with potential prey (i.e., their larvae kill aphids) they were less commonly flying around cucumber plants since aphid numbers were generally lower in partridge pea and weedy plots.
Ladybeetle adult abundance was influenced by treatment (Anova: F(2, 176.284) = 3.786, p = 0.025). Specifically, they were more abundant in partridge pea treatments versus controls (p = 0.0375), but not between weedy treatments and controls (p =0.099) or partridge pea and weedy treatments (p = 0.908). Temperature had a significant positive correlation with ladybeetles (Anova: F(1, 40.054) = 31.825, p < 0.001, partial r-squared = 0.179), but humidity did not (Anova: F(1, 8.348) = 0.100, p = 0.760)(Table S15 and S16)(Fig. S5). This result makes sense, as ladybeetles are known voracious generalist predators, which also seek carbohydrates in the form of nectar. I personally saw ladybeetle adults feeding directly from EFNs of our partridge peas (something I did not observe in the field with any other group besides ants and wasps).
Rove beetles (Family: Staphylinidae) did not vary in cucumber plots by treatment (Chi-squared = 0.335, DF = 2, p = 0.846) or humidity (Chi-squared = 0.690, DF = 1, p = 0.406, but their abundance was positively correlated with mean temperature (Chi-squared = 13.425, DF = 1, p = 0.0003, (Table S17). Overall, rove beetles were not very numerous on these sticky cards, and were mostly a single small species. Also, as ground-dwellers, these beetles are more commonly found in pitfall traps (however not very common in ours, and were thus not analyzed from our pitfalls).
Generalist wasp abundance (Family: Vespidae) was significantly higher in partridge pea plots than both control and weedy ones (Chi-squared = 83.238, DF = 2, p < 0.0001), but their abundance was not correlated with temperature (Chi-squared = 0.105, DF = 1, p = 0.746) or humidity (Chi-squared = 0.115, DF = 1, p = 0.735)(Table S18 and S19)(Fig. S5). These results align similarly with our ladybeetle data. Vespid wasps like yellowjackets and paper wasps, like ladybeetles, are predatory generalists which feed on nectar. Overall, the significant positive effect of temperature on many of these insect groups’ abundance is not too surprising, as these collections were made progressively throughout the summer, and higher temperatures allow for higher activity of small exotherms like insects. However, it is also important to note that the most of the r-squared values from analyses that were significant were fairly low, suggesting that temperature did not explain much of the variation seen in their respective datasets.
The compositions of the above insect groups were found to vary significantly among treatments after analysis via Permanova (F(2, 190) = 1.829, R2 = 0.019, p = 0.04)(Table S20)(Fig. S6). However, post-hoc pairwise comparisons yielded no significant p-values, with the closest to significance being partridge pea versus weedy (0.06)(Table S21). These results are similar to the above comparison of ant species composition among treatments. While there was a significant main effect and near-significant post-hoc result between two groups, the polygons are mostly overlapping, suggesting that they are mostly similar in composition of the groups that were identified from sticky cards.
Objective 2
Yield
There was a significant difference in total cucumber yield, with both partridge pea and control plots being higher than weedy ones (Anova: F(2, 636.12) = 4.165, p = 0.016 (Table S22 and S23)(Fig. S7). Marketable cucumber yield (Anova: F(2, 611) = 3.164, p = 0.043) (Table S24 and S25)(Fig. S8), the number of total fruits (Anova: F(2, 390.01) = 8.311, 0.0003)(Table S26 and S27)(Fig. S9), and number of marketable fruits (Anova: F(2, 339.58) = 12.488, p < 0.0001)(Table S28 and S29)(Fig. S10) were also higher in partridge pea and control plots versus weedy plots. When calculating the hypothetical profit if our produce was sold and then adjusting for costs incurred from growing partridge pea, the treatments followed the same pattern as above: with profit from both partridge pea and control being significantly higher than weedy plots (Anova: F(2, 42.39) = 6.345, p =0.004) (Table S30 and S31)(Fig. S11). While I expected the partridge pea-treated plots to produce the most cucumber over both weedy and control plots, these results still help support the concept that the inclusion of these EFN-producing plants provide benefits when growing cucumber. Cucumber growing next to partridge were importantly not significantly lower than the control plots. The fact that they were on par with the controls show that any negative impacts that may be introduced by the addition of partridge pea (i.e., resource competition, shading) were outweighed by the benefits (i.e., increased abundances of generalist predators, potentially increased pollination services).
Leaf damage and disease
Overall, there was very little leaf damage across all treatments, making our dataset very right skewed. I ran a nonparametric test to determine differences in leaf damage across treatment. There were no significant differences after conducting a Kruskal-Wallis Rank-Sum test (Chi-squared = 5.649, DF = 2, p = 0.06). The proportion diseased cucumber leaves did not vary among the three treatments after conducting a chi-squared contingency test (Chi-squared = 0.935, DF = 2, p-value = 0.627). Due to low damage this season, any differences were likely caused by a few leaves that had > 10% damage.
Objective 3
Companion planting in cucumber versus zucchini
Trapped ant species richness varied by the interaction between crop type (i.e., zucchini vs. cucumber) and treatment (Anova: F(1, 102.829) = 4.067, p = 0.046)(Table S32). However, a post-hoc analysis revealed no significant individual comparisons (Table S33). Ant richness was significantly higher on zucchini overall (β = -0.8043, SE = 0.340, DF = 93.962, t = -2.362, p = 0.020) but not by treatment (β = -0.563, SE = 0.302, DF = 60.484, t = -1.866, p = 0.067) when analyzed as fixed effects alone. There was also no significant correlation with soil compaction and ant species richness (Anova: F(1, 116.984) = 4.292, p 0.041) (Table S32). These results reflect those of the above analyses from the 2025 cucumber dataset, with no significant differences between treatment type. However the findings that there were more ant species overall on zucchini are interesting, and possibly due to the presence of EFNs on zucchini supporting more ant species.
Ant community composition varied significantly by the interaction of crop type and treatment after conducting a Permanova (F(3, 116) =1.835, R2 = 0.047, p = 0.024)(Table S34). After conducting a post-hoc analysis, the only significant difference was between control and partridge pea cucumber plots (p = 0.006)(Table S35). When visualised, there was some overlap between these two groups, but also a relatively equal portion of these polygons did not overlap vertically on the NMDS2 axis (Fig. S12). This may suggest that the collection of ant species trapped in cucumber next to partridge pea is different from those trapped in control plots, with some common species linking the two. However this result does not align with the above NMDS analysis when including the entire dataset from 2025. Only the single site (Spencer, MA) was included in this analysis, as zucchini was only grown there in 2024. Therefore, the inclusion of the second site when analyzing ant species composition likely has an impact on differences seen. In general, my experience working on small organic farms has shown me that every farm is unique in terms of its insect community. These differences are caused by the varying land management practices at each farm, land use history, produce and animals being raised, and proximity to land features like lakes, forests, or cities.
When examining striped cucumber beetle abundance from sticky cards with zucchini data from 2024 included, there was no effect of the interaction of treatment or plant type on striped cucumber beetle abundance (Chi-squared = 0.202, DF = 1, p = 0.653). When examining these factors individually, there were overall significantly more striped cucumber beetles on zucchini than cucumber (β = 0.994, SE = 0.216, z = 4.60, p < 0.0001), but there were no significant differences between partridge pea and control plots (β = -0.431, SE =0.246, z = -1.755, p = 0.079). There was no correlation between striped cucumber beetle abundance and soil compaction (β = 0.210, SE = 0.267, z = 0.787, p = 0.432). Table S36). While no influence of treatment was present, the difference in striped cucumber beetles between the two plants was not very surprising, as I remember seeing many more striped cucumber beetles in the zucchini plots in 2024 compared to the subsequent year when I grew cucumber at that same site. Our model initially included sampling date and year as random effects, but were removed after they had near zero effect sizes, but there could have been more underlying variables that I did not measure like the inherent differences in phenology of these two crops, or the difference in plant volume between the two (zucchini in general are much taller and wider). However, it could also be true that in this area of Massachusetts striped cucumber beetles are more prone to be attracted to and feed on zucchini versus cucumber.
Zucchini marketable weight did not significantly vary between control and partridge pea plots in 2024 (Table S37). (Anova: F(1, 398.13) = 1.253, p = 0.264)(Table S37). The number of marketable zucchini fruits also did not vary (Anova: F(1, 119.77) = 1.861, p = 0.175)(Table S38). These two plants inherently have different outputs in fruit production and weight. Thus, I could not directly compare their yield in a single analysis. These results, in tandem with those of the cucumber harvest data, show that the productivity of both zucchini and cucumber were not significantly increased or reduced by the presence of partridge pea.
Literature Cited
1. Jones, I. M., Koptur, S. & von Wettberg, E. J. The use of extrafloral nectar in pest management: overcoming context dependence. J Appl Ecol 54, 489–499 (2017).
2. Bentley, B. L. Extrafloral Nectaries and Protection by Pugnacious Bodyguards. Annu. Rev. Ecol. Syst. 8, 407–427 (1977).
3. Arimura, G., Kost, C. & Boland, W. Herbivore-induced, indirect plant defences. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 1734, 91–111 (2005).
4. Koptur, S. Extrafloral Nectary-Mediated Interactions Between Insects and Plants. 50 (1992).
5. Bronstein, J. L. The Contribution of Ant-Plant Protection Studies to Our Understanding of Mutualism1. Biotropica 30, 150–161 (1998).
6. Morris, J. B. Showy partridge pea [Chamaecrista fasciculata (Michx.) Greene] with potential for cultivation as a multi-functional species in the United States. Genet Resour Crop Evol 59, 1577–1581 (2012).
7. Karasevicz, D. & Crist, C. Bacterial Wilt of Cucurbits. 1–2 https://chemung.cce.cornell.edu/resources/bacterial-wilt-of-cucurbits (1996).
8. Rotolo, C. et al. Use of biocontrol agents and botanicals in integrated management of Botrytis cinerea in table grape vineyards. Pest Management Science 74, 715–725 (2018).
9. Sharma, N. & Singhvi, D. R. Consumers perception and Behaviour towards organic food: A systematic review of literature. J Pharmacogn Phytochem 7, 2152–2155 (2018).
10. Ware, G. W. Effects of pesticides on nontarget organisms. in Residue Reviews (eds Gunther, F. A. & Gunther, J. D.) 173–201 (Springer, New York, NY, 1980). doi:10.1007/978-1-4612-6107-0_9.
11. Howarth. F. Environmental Impacts of Classical Biological Control. Annual Review of Entomology. 27. 485-509 (1992).
12. Koch, R. L. The multicolored Asian lady beetle, Harmonia axyridis: A review of its biology, uses in biological control, and non-target impacts. Journal of Insect Science 16 (2003).
13. Mendelsohn, R. The Impact of Climate Change on Agriculture in Developing Countries. Journal of Natural Resources Policy Research 1, 5–19 (2009).
14. Gahukar, R. T. Entomophagy and human food security. Int. J. Trop. Insect Sci. 31, 129–144 (2011).
15. Bugg, R. L. & Waddington, C. Using cover crops to manage arthropod pests of orchards: A review. Agriculture, Ecosystems & Environment 50, 11–28 (1994).
16. Herz, A. et al. Managing Floral Resources in Apple Orchards for Pest Control: Ideas, Experiences and Future Directions. Insects 10, 247 (2019).
17. Haber, A. I. et al. Striped cucumber Beetle and Western Striped Cucumber Beetle (Coleoptera: Chrysomelidae). Journal of Integrated Pest Management 12, 1 (2021).
18. Anjos, D. V. et al. The effects of ants on pest control: a meta-analysis. Proc. R. Soc. B. 289, 20221316 (2022).
19. Campbell, J. W., Irvin, A., Irvin, H., Stanley-Stahr, C. & Ellis, J. D. Insect visitors to flowering buckwheat, Fagopyrum esculentum (Polygonales: Polygonaceae), in north–central Florida. The Florida Entomologist 99, 264–268 (2016).
20. Schifani, E. et al. New Tools for Conservation Biological Control: Testing Ant-Attracting Artificial Nectaries to Employ Ants as Plant Defenders. Insects 11, 129 (2020).
21. Agarwal, V. M. & Rastogi, N. Deterrent effect of a guild of extrafloral nectary-visiting ant species on Raphidopalpafoveicollis , a major insect pest of sponge gourd, Luffacylindrica. Entomologia Experimentalis et Applicata 128, 303–311 (2008).
22. Phillips, B. W. & Gardiner, M. M. Does local habitat management or large-scale landscape composition alter the biocontrol services provided to pumpkin agroecosystems? Biological Control 92, 181–194 (2016).
23. R Core Team. R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, 2025).
24. Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software 67, 1–48 (2015).
25. Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. R package version 0.4.7 http://florianhartig.github.io/DHARMa/ (2024).
26. Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.11.2-8 https://doi.org/10.32614/CRAN.package.emmeans (2025) doi:10.32614/CRAN.package.emmeans.
27. Oksanen, J. vegan: Community Ecology Package. R package version 2.7-2 https://doi.org/10.32614/CRAN.package.vegan (2025) doi:10.32614/CRAN.package.vegan.
Conclusions
Ants are the most important mutualist groups involved in EFN-insect interactions, but the number of ant species and their composition was not heavily influenced by the addition of partridge pea. However, the abundance of other important EFN-seeking generalists like wasps and lady beetles were significantly higher in partridge pea plots than controls. These results, coupled by the fact that aphid counts were lower on sticky cards in control plots, help assert the notion that the inclusion of extrafloral resources can provide some benefit to the main crops being grown. The abundance of striped cucumber beetles, which are the most economically-important insect pest on cucurbits in this region, were not significantly influenced by the companion plant treatments. It is also important to note that striped cucumber beetle numbers were generally low at both sites this year, with a total of 137 being trapped on sticky cards, and a total of 32 Chrysomelid beetles were collected from pitfalls.
Harvest, leaf damage, and disease results show that partridge pea did not have a significant influence on the productivity or health of our cucumber. However, partridge pea was also not significantly detrimental towards cucumber in any of these aspects, unlike the weedy treatment which was significantly lower than both partridge pea and control plots in terms of production. Importantly, costs associated with including partridge pea as a companion plant did not offset these findings. Additionally, partridge pea similarly influenced zucchini: fruit productivity was neither significantly increased or reduced with its inclusion as a companion plant. Striped cucumber beetles were more abundant on zucchini sticky cards, but these differences are likely due to the plants themselves than any treatment effects. I did not assess the influence of partridge pea as a nitrogen-fixing legume on soil quality as it was out of the scope of my dissertation. It is important to note, that uprooting these plants at the end of the field season showed how incredibly proficient they were in forming rhizobia nodules. While the effects of nitrogen replenishment wouldn’t be seen until the following seasons, this is further evidence that these plants are a good option as a companion plant or cover crop. With these results, I can confidently state that partridge pea is a good candidate as a companion plant for organic farmers seeking to provide resources to attract and maintain beneficial insects from the surrounding landscape. Importantly, more studies like this are needed to further understand the synergistic effects that EFN-producing plants provide when planted on farms in their native range.
Education & outreach activities and participation summary
Participation summary:
I conducted two types of outreach activities throughout the summer of 2025: 1) informational booths at local farmer's markets and 2) insect collection and identification workshops at local parks with K-12 students.
Farmer's markets
I reached out to a representative of the Xerces Society and was able to get pamphlets (see attached pamphlet PDFs) sent to us that were tailored toward beneficial insect conservation in the Northeast US. We then set up an informational table (Fig. 9) at four separate farmer's market dates, 3 in Worcester and one in Grafton, MA. At these events, Me and the undergraduate students in our lab educated visitors on what we do at Clark, talked about the importance of insect diversity in agriculture, and described the science behind the project that I outlined in this grant.
The pamphlets we handed out described how hobbyists and professional farmers can manage the landscape around their plantings by choosing companion plants that complement their specific crops, as well as how to identify native beneficial insects. The in-person activities I have described here will also benefit the undergraduate students working for us during the summer, as they will encourage them to participate and build on their skills in public outreach as young scientists. Additionally, I will share yield and insect diversity results with local farmers as I gather and analyze our data. I will present the summaries of our results at one of the local symposia held by the Northeast Organic Farming Association (NOFA), as well as publish findings in one of their newsletters free to the public.
- 22-014_01_Soil-Bioindicators-Guide_web-screen
- 08-005_02_XercesSoc_Farming-for-Pest-Management-brochure_web
- 12-020_03_BumbleBeeConservation_web-print
- 08-006_01_XercesSoc_Farming-for-Pollinators-brochure
- 18-014_02_Natural-Nesting-Overwintering-FS_web
- 15-031_04_HAG_FarmsAgLandscapes_web 22-026_01_NPPBI—Northeast_web
- 16-044_02_MNPL_Northeast_web-print
K-12 student outreach
I worked alongside leaders at Recreation Worcester, a free youth summer program run by our city, to design a curriculum aimed at teaching kids about the importance of native insects and identify the major groups of arthropods they may find while playing outdoors. I created a booklet going over these insect groups, which we printed and handed out to the kids (see attached "Bugs in your backyard" PDF). These events took place at 3 public parks, ranging from ~20-30 kids and a handful of chaperones participating at each. For the activities, we spent ~2-3 hours at each park collecting insects from trees and vegetation with sweep nets, placing them in collection vials, and then passing them around to the kids while educating them about their ecological importance and what interactions they may have with humans. The educators we partnered with thought last year’s outreach event was successful and provided an educational benefit to the kids who participated, and they invited us to join in their 2026 season. We will be doing similar activities with them in August this year.
Fig. 8: Table setup at one of the Worcester public farmers market events

Project Outcomes
These results are promising, in that I provide evidence that partridge pea may be a viable companion plant in organic cucurbit systems. Partridge pea has the potential to increase native pollinator services, provide valuable carbohydrate resources and refuge for generalist predators, and replenish nitrogen availability (as partridge pea is a legume) in their fields. I suggest that future conservation biocontrol field experiments like mine be conducted on a variety of different organically-grown crops. Even if direct benefits to yield are not directly seen within the first season, the advantages of including companion plants on a farm over multiple years will likely accumulate over time.
The influence that extrafloral nectar has in agroecosystems and the beneficial predatory arthropods that are attracted to EFNs is still heavily understudied compared to the classic flower-pollinator system. Maintaining resource-provisioning plants over consecutive growing seasons on a farm will help support a healthy population of beneficial native organisms like pollinators, predators, and nitrogen fixing microbes that have spent millions of years forming mutualisms with native plants, such as partridge pea, in New England. The experimental design and outcomes from this project will make a great baseline for future studies that aim to answer similar questions in different planting systems and different regions of the world.
Learning reflections
The most important outcomes from this project is that 1) the inclusion of partridge pea did not significantly reduce the productivity of cucumber and 2) there were significant increases seen in generalist predators (lady beetles and wasps) and reductions in aphid numbers when comparing partridge pea plots to controls. Partridge pea produces a high quantity of extrafloral nectar throughout the growing season, which is very attractive to ant workers. Additionally, the flowers of partridge pea seem to be a good resource for attracting native bees. During our sampling days our partridge peas had very high bee activity, with dozens of bumblebees, solitary bees, and predatory wasps foraging in each PP-treated block at any given time. Knowing that partridge pea does not significantly reduce cucumber yield like the weedy fallow treatment did, it seems like this plant would be a useful addition for farmers looking to enhance the ecosystem services provided by arthropods from the surrounding landscape in their farm.
In general, I learned a great deal about sustainable agriculture while working on this project. From two summers of tilling, hoeing, mulching, planting, irrigation/repairing drip tape, adding/removing row covers, staying on top of weeding, mowing between rows, and harvesting - I have gained much respect for the farmers that throw aside the conveniences of industrial agriculture. I also learned how much organic farmers care about the health of the land that they manage, and how important it is for them to stay on top of modern research and methods for growing food in a manner that is safe for the surrounding landscape. Thus, the results gained from this study will greatly benefit such farmers looking for novel methods in sustainable food production.
Future research/career directions
For future directions I plan to look into the specific interactions among organisms that take place in these cropping systems. Other components of my dissertation focus on 1) the relationships and fitness impacts between nectar microbes, plants, and ants in agroecosystems, and 2) how ant cuticular hydrocarbons (i.e., the scent trails they leave behind while foraging) influence the behavior of herbivorous insects. In terms of my career, I am interested in continuing in academia and plan to pursue a postdoc position at a university studying plant-insect-microbe interactions.
Assessment of study and outreach
My study was the product of multiple years of planning, and I feel that its design is very sound. Our outreach work was very successful in my opinion, as we were able to engage and inform our target audiences. However, one challenge that I did not expect during our outreach was attempting to get the older kids involved in the insect collection activities when working at parks with Recreation Worcester. The younger elementary/junior high students we worked with were much more interested in our activities, generally participated more, and asked thoughtful questions. My assumption from this is that the older students were more interested in taking the day to hang out with their friends, and saw our workshop as more busy work instead of a fun activity.
Our field work and outreach could not have been completed without the help of the undergraduate students this summer, which were compensated using the funds from this SARE grant. As a PhD student, I do not make much money, especially since we do not get paid during the summer when not teaching. This grant supported me and allowed me to put all of my effort into the work I conducted this summer. However, the limits of a 5-6 year dissertation mean that the number of seasons that such a study can be conducted are limited. From the knowledge and experience from other agricultural researchers, I know that the effects and applicability of results to real-world scenarios of an agroecological study system is highly dependent on location, resources, and time.
Time is an incredibly important factor that influences the effectiveness of something like a cultural/conservation biocontrol method or companion planting treatment, as it may take multiple seasons of the same planting regime to build up beneficial invertebrate communities in the soil and surrounding environment. Insects can also be highly variable from farm to farm in a given geographic region, especially between organic farms whose managers have their own areas of expertise and preferred pest control practices. Therefore, being able to include more sites in this study would have also been useful.
Further study
While out of the scope of my own dissertation, it would be interesting to see what influences partridge pea may have on the community of soil microbes and soil nutritional quality, especially since they form mutualisms with nitrogen fixing bacteria. Partridge pea are a native legume, and may be more suited to acquiring soil mutualists in their native range of the Northeast US compared to other non-native legumes grown here like garden peas and beans. Moreover, these mutualisms may change the chemistry of the soil surrounding the crops that they are planted next to. Future work should examine the influences that such alterations may have on the health and nutritional quality of the focal crop being produced.