Final Report for LS10-229
Project Information
We determined the effects of integrated crop-livestock (ICL) systems on soil microbial community composition, C sequestration potential, and global warming potential (GWP) in the semiarid Southern High Plains (SHP). ICLs, especially perennial components, had multiple positive effects on ecosystem health including increased C sequestration potential, macroaggregate formation (increased stability), & enhanced nutrient cycling. These ecosystem benefits were associated with higher fungal diversity and specifically the relative abundance of arbuscular mycorrhizal fungi (AMF) and Gram negative (GM-) bacteria. Among ICLs, the GWP of the dryland ICL was estimated to be near neutral, whereas the deficit-irrigated ICL was a net C-source.
Our hypothesis is that the integrated crop and livestock systems will support a more diverse microbial community, sequester more C in long-term pools and produce lower emissions of GHGs, compared to the continuous cotton system. Furthermore, we believe that despite water limitations these systems in a semi-arid environment will prove to be a valuable player in the global search for C sequestering systems. To achieve this goal we will use existing SARE-funded long-term replicated, integrated crop and livestock systems (irrigated and non-irrigated) and three on-farm producer crop and livestock systems that are a part of the TAWC.
Specific objectives are to:
- Measure the amount of C stored in active, intermediate and passive SOM pools using a detailed physical fractionation method, which provides eight different soil aggregate fractions.
- Evaluate greenhouse gas fluxes (CO2 and N2O) to calculate global warming potential within each system.
- Characterize soil microbial community structure using fatty acid methyl ester (FAME) profiling and microbial biomass C (MBC) including bacterial diversity in whole soil and different soil aggregate fractions using a novel molecular biological tool (i.e., pyrosequencing).
- Translate results from Objectives 1 through 3 into practices incorporated in agriculture in the THP and similar ecosystems. Specifically, we will increase producer and consultant awareness regarding the direct and indirect positive effects for managing agricultural lands to achieve enhanced soil functioning.
Our long-term objective is to use the data acquired under the proposed research to guide policy-makers and agricultural and land managers to make the best ecological and socio-economical-supported decisions possible. In doing so, they will meet the increased agricultural demands for food and fiber while sustaining natural resources to preserve and enhance local communities. Information we acquire will supplement past and concurrent data, including an additional Ph.D. research project (in progress) that is evaluating energy use, efficiency and economics in the proposed sites. These established long-term replicated integrated research sites as well as access to grower-managed farms with known histories and similar systems, provides an unparalleled opportunity enabling us to achieve such an important yet complex goal. Subsequent funding will enable us to provide detailed assessment of C sequestration potential and soil microbial biodiversity estimates in these semiarid agroecosystems; systems which may serve as models for sustainable agricultural production across this agriculturally, socially, and economically important eco-region.
The purpose of this project is to assess the capacity of long-term, integrated crop and livestock systems in the semi-arid Texas High Plains (THP) to enhance soil quality and functioning by evaluating soil microbial diversity and its link to soil processes including carbon (C) sequestration and greenhouse gas (GHG) fluxes. Although, there is a strong interest in optimizing agricultural management to sequester C in soils to offset global climate change, this has not been a major priority to producers in the THP. Within Texas, the Southern High Plains (SHP) region encompasses more than 11.6 million acres (4.7 million ha) of cropland and over 30% of the U.S. cotton (Gossypium hirsutum L.) crop is produced in this region, primarily in monocultures with conventional tillage. Cotton produces far less residue per hectare than do other major crops (Lal, 2004) and intensive tillage operations have been associated with accelerated decomposition of soil organic matter (SOM) and subsequent loss of C to the atmosphere as CO2. Cultivated intensively since about 1940, irrigated agricultural production in the THP has been at the expense of high water use that has exceeded the recharge potential of the Ogallala aquifer in this low rainfall region. This practice has provided high economical profit but has come at the expense of soil exposure during winter fallow periods leading to increased wind soil erosion and significant reductions in SOM content. Today, regional challenges imposed by impending loss of the Ogallala aquifer for irrigation, low fertility soils, high soil erodibility, and stressful climatic conditions have prompted producers to reconsider their options as past successes are at risk.
While the potential to sequester C is strongly associated with water, this semi-arid region is positioned to play an important role because degraded soils whose SOM reserves have been depleted due to soil mismanagement (such as long-term continuous cropping) hold the largest potential for soil organic C SOC sequestration (Lal, 2009). Furthermore, many acres are available in this vast landscape to contribute to sequestering C and with impending scarcity of water for irrigation, cropland is increasingly being returned to grass production. Perennial forages protect fragile soils, convert atmospheric C to SOM, and reduce water and fertilizer requirements compared with current cropping practices. Pastures have the potential to store significant amounts of SOC (Cerri et al., 2003) because their dense and deep root systems contribute to aggregate formation and the inclusion of C within, which is less susceptible to oxidation and eventual loss to the atmosphere (Cambardella and Elliott, 1993, 1994). Therefore, perennial forage systems may provide an important means for C storage in these semiarid lands where degradation of SOM exceeds the rate of stabilized C formation due to the production of low residues with the intensively tilled cotton monoculture system (Acosta-Martínez et al., 2003).
Carbon is stored in SOM within various functional pools with certain turnover rates ranging from labile (readily decomposable; fast turnover) to very recalcitrant (resistant to degradation; slow turnover). According to Lal and Follett (2009), the SOC pool can be increased when C inputs exceed C outputs through land use conversion and/or adoptions of conservation-effective measures that reduce the loss of nutrients and water and increases biomass production. Studies have reported that macroaggregate formation in less disturbed systems provides an important mechanism for the protection of SOC that may otherwise be oxidized with aggregate breakage (Six et al., 1998). In order to assess management impacts on SOM storage, the amount of C incorporated into the various pools must be quantified and characterized to better understand processes of C sequestration for the THP. Researchers have developed physical (size and density) fractionation methods to quantify the magnitude of soil C being incorporated in these fractions as a result from land use changes or climate change scenarios. The resulting soil aggregates are classified in terms of size and relative stability and thus, C associated with soil pools which protect it from decomposition are considered to be an indication of long-term SOC sequestration. For example, Six et al. (2000), showed that increased SOC under no-till compared with conventional tillage management is the result of stabilization of C within microaggregates and that this component is very stable. These findings were only made possible via a fractionation scheme that can differentiate the 'free' microaggregates from those already stabilized within macroaggregates (Six et al., 1999); fractions which are not isolated with whole SOM characterization. Of the various fractions isolated using the method as described by Six et al. (2000), the microaggregates within the macroaggregates have been proposed as an early indicator for potential SOC sequestration (Denef et al., 2007; Kong et al., 2005).
Sustainable Agriculture Relevance
Agriculture as it is currently practiced today in the THP is not sustainable. It is based on a water resource that will not be there in the future as it has been in the past. Additional challenges facing agricultural production in this region include soils of low fertility, high erosion potential, soil salinity issues, and a harsh climate (high summer temperatures, low humidity and rainfall, high winds, and damaging hail). The THP is representative of the much larger Southern High Plains. Furthermore, it is positioned to provide a major site in the needed network of long-term agricultural ecosystem sites where a greater understanding of mechanisms and processes governing soil ecology and C dynamics can be elucidated and then be put to appropriate use. Managing lands for C sequestration should help offset increasing atmospheric CO2 concentrations while improving site fertility, agricultural sustainability, soil biodiversity, and water conservation. More information is needed, however, on the effects of alternative agricultural systems on C cycling and the potential to develop integrated cropping and livestock systems to optimize C capture while meeting production, economic, and environmental goals. In a broader perspective, our success will be measured by the future of this region where 40% of the economy is based on agriculture. Our research on these integrated systems will address an important aspect of systems sustainability by focusing on ecosystem services performed by soil systems such as C sequestration, enhanced microbial diversity, and reduced GHG emissions.
Currently, there is a movement by policymakers to allow industries (such as factories) who exceed their GHG emissions cap to pay incentives to encourage farmers to adopt practices to off-set C emitted and that curb GHG. Our results from the proposed project will provide a framework to help guide management decisions to accomplish goals of profitability and the environment. Solutions found here can provide the principles that other ecoregions can follow. To find economically and environmentally sustainable practices, long-term systems-based agroecosystem research is required in site-specific ecoregions and must be conducted by proven interdisciplinary teams of researchers, producers, and others representing the wide array of stakeholders and policy makers. We have in place already the capacity to continue to conduct such long-term research in one of the most important, but challenged agricultural ecosystems in the U.S. Such a network of sites is critical to understanding agricultural systems so that the mechanisms and processes governing specific processes such as soil ecology and C dynamics can be elucidated and this information can then be put to appropriate use. This is the foundation of a sustainable US food and agricultural industry and a sound national economy. Understanding the mechanisms underlying C sequestration in agroecosystems is critically needed to ensure management decisions accomplish objectives. Carbon sequestration in the THP is challenging due to our warm, semi-arid environment, decreasing water resources and due to the low residue return of the main crop, cotton. Preliminary research in the irrigated systems in the original SARE-funded project (LS97-82) has shown that integrated a crop/forage/cattle production system conserved water, nutrients and SOC and increased microbial activities (Acosta-Martinez et al., 2004; Allen et al., 2008). However, the mechanisms underlying these results remain undetermined and no such research has been conducted in the non-irrigated and irrigated systems (LS02-131) in the THP.
We will ensure that we are not substituting one ecological problem (e.g. reduced SOC storage) with another (e.g. increased energy demands) by coupling our results with, for example, a concurrent Ph.D. project whereby a complete energy budget is being conducted. To our knowledge, this information is not available for integrated crop-livestock systems under non-irrigated and irrigated management. With this detailed research approach, we will improve the ecological impact assessment for these semi-arid agroecosystems by evaluating the biogeochemical mechanisms for C storage and assessing management practices which result in reduced GHG fluxes and maintain or enhance biological diversity. The end result will better equip agricultural and land managers to make the best ecological and socio-economical-supported decisions possible.
Partitioning of residue-C between CO2 via respiration and storage in semi-permanent soil-C pools is largely controlled by the composition and activities of the soil microbial community. Microbial communities comprise the biologically most active fraction of SOM (Smith and Paul, 1990; Franzluebbers et al., 1997), and aggregate formation results from their decomposition products or by mechanical binding of soil by fungal growth (Lynch and Bragg, 1985). Thus, fungal and bacteria play different roles in soil aggregate formation. However, despite the important role of soil microbial communities in C sequestration and the formation of macroaggregates and microaggregates, attempts to determine microbial community distribution within various size classes has received very little attention (Mummey et al., 2006).
Fatty acid profiling is a commonly used approach to determine broad-level microbial community characterization. It can distinguish among general taxonomic groups such as fungi, protozoa, and bacterial groups such as Actinomycetes, Gram-positive and Gram-negative bacteria, but it cannot provide information at the species level. Molecular biological techniques based on DNA sequencing are providing a step further into characterization of the microbial community structure of soil. However, traditional cloning techniques have been considered impractical to adequately measure the vast microbial diversity of soil (Gans et al., 2005). Recent advances in sequencing technology, such as the development of pyrosequencing, have lead to fast and massive sequencing approaches (i.e., 400-600 MB output per run in a day), which can provide between 4500 to 52000 unique sequences or operational taxonomic units (OTUs; roughly corresponding to the species level) in soils. This output is between 10 to 100 times higher compared to what can be obtained (i.e., about 335 OTUs) using traditional cloning techniques (Roesch et al., 2007; Acosta-Martinez et al., 2008). Moreover, pyrosequencing is based on a 'sequence-by-synthesis' method that overcomes cloning bias and labor-intensive tasks required by the traditional approach with the Sanger method (Cardenas and Tiedje, 2008). Thus, pyrosequencing is a promising new tool that will expand our understanding of the microbial community structure of soils quickly and more comprehensively than other molecular approaches currently in use (Roesch et al., 2007; Dowd et al., 2008a; Acosta-Martinez et al., 2008).
Opportunity and solutions: We have a unique research opportunity to assess the capacity of integrated crop and livestock systems to enhance soil microbial diversity and its link to C sequestration potential with long-term fieldscale plots already established in the semi-arid THP. In 1997, with funding largely from the USDA-SARE program (LS97-82), researchers at Texas Tech University established replicated long-term field-scale comparisons of an irrigated cotton monoculture with an integrated, irrigated, 3-paddock cotton/forage/beef cattle system at the Texas Tech Experimental Research Farm in New Deal, TX. Initial successes led to further USDA-SARE funding in 2002 in which a non-irrigated, 3-paddock cotton/forage/beef cattle system and an irrigated all forage/beef cattle system were added on an additional 103.7 acres (42 ha). This ongoing research and education platform now includes nearly 148 acres (60 ha) in four replicated systems to compare total water applied (irrigation plus precipitation), economics, energy balance, and total productivity of plants and animals and was selected by the USDA as a national model of successful research. Additionally, an array of 27 on-farm producer-managed crop and/or livestock systems was added in 2005 with subsequent Texas state funding. This research platform, totaling over 4000 acres (1600 ha), provides us with an unparalleled opportunity to answer specific questions regarding soil C dynamics previously not investigated within these agroecosystems. The investigators are actively engaged in research relevant to this topic. Specifically, team members have published on the early changes in whole soil C content and on microbial communities and activities as affected by SARE-funded systems comparisons, which were under the irrigated three paddock cotton/forage/beef cattle system and the monoculture cotton (Allen et al., 2005; 2008; Acosta-Martinez et al., 2004; 2008). Results from the overall project has shown that the integrated system used 25% less irrigation water, 40% less nitrogen fertilizer, had increased SOC and microbial activity, a lower soil erosion potential, and was comparable in profitability with the cotton monoculture at yield levels typical of the region (Allen et al., 2005; 2008). However, research conducted on the effects of the non-irrigated systems begun in 2002 is limited and no study to date has characterized the various fractions of SOM under the proposed management systems. Additionally, we now have the opportunity to investigate these soil properties under production conditions on producer sites within the state-funded (TAWC, 2009) demonstration project. Moreover, we know of no study that has attempted to assess how the SOM fractions interact with soil biota at the level of detail we intend to achieve. There is a great need for detailed information on the effects of these agricultural systems on C cycling and the potential to develop integrated systems to optimize C capture and sustain biodiversity while meeting production, economic, and environmental goals. Therefore, our overall goal is to find sustainable agroecosystems for the THP that can be applicable to other semiarid regions with fragile soils and water availability problems, by linking soil microbial communities to changes in ecosystem functioning (i.e., C sequestration and reduced GHG fluxes). This research is needed to assist land managers and researchers to identify viable agricultural systems, and to provide the education to extend this knowledge into practice.
Cooperators
Research
Study sites and management details
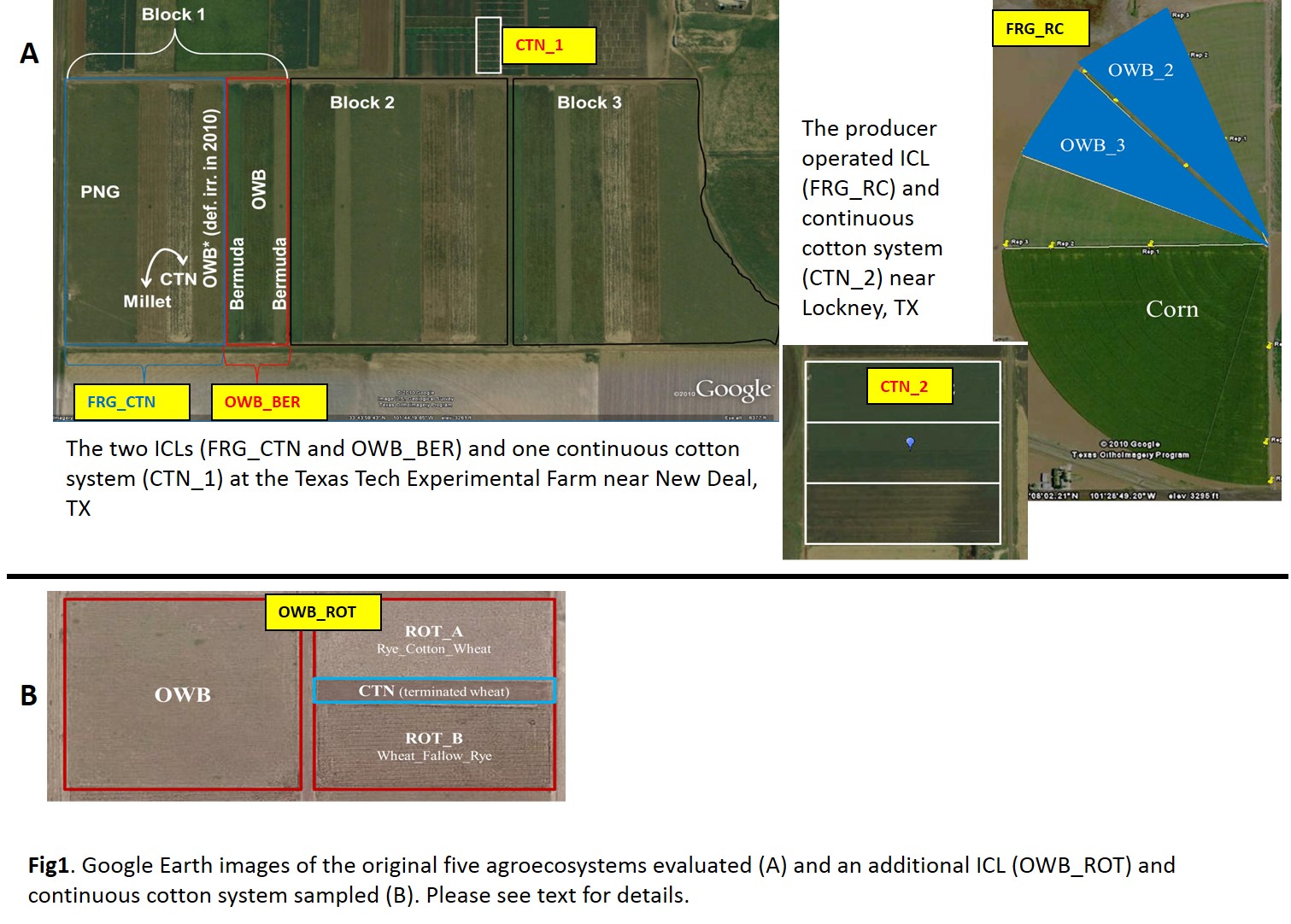
A total of five agroecosystems were selected to represent continuous cotton (CTN), the dominant form of agricultural production in this region, and integrated crop-livestock (ICL) agroecosystems, a potential alternative management practice (Fig 1a). All agroecosystems were located in the Southern High Plains region of northwest Texas in a semiarid climate with an average annual precipitation of 466 mm and average evapotranspiration (ET) of 1500 mm. The average daily temperature ranges between 7.9 and 22.9 ?C. The coolest month is January with an average low temperature of −4.4 ?C, and the warmest month is July with an average high temperature of 33.3 ?C. Between January and just prior to sampling in July 2010, the study site received 40.6 cm of precipitation. Three of the agroecosystems (2 ICLs and 1 CTN) were located on the Texas Tech Experimental Farm six miles east of New Deal, TX (N33.734040°, W101.738706°). The remaining two agroecosystems (1 ICL and 1 CTN) were producer-operated and participants in the Texas Alliance for Water Conservation (TAWC) program and were located near the town of Lockney, TX (N34.124518°, W101.441552°), 50 km northeast of the Texas Tech Experimental Farm.
- The first ICL (FRG_CTN; 10 ha), initiated in 2004, was comprised of three non-irrigated paddocks and one irrigated paddock. The non-irrigated paddocks included one paddock of perennial native grasses (PNG; 4.5 ha) dominated by blue grama [Bouteloua gracilis (Willd. ex Kunth) Lag. ex Griffiths], sideoats grama [Bouteloua curtipendula (Michx.) Torr.], buffalograss [Buchloe dactyloides (Nutt.) J.T. Columbus], and green sprangletop [Leptochloa dubia (Kunth) Nees]. The two remaining paddocks (1.7 ha each) consisted of annual crop components cotton (R-cotton – FiberMax 905F) and foxtail millet (R-millet – Setaria italic [L.] P. Beauv.). The two annual components were managed under conventional tillage and planted in rotation such that both crops were present each year. The irrigated paddock contained ‘WW-B. Dahl’ old world bluestem (OWB1) [Bothriochloa bladhii (Retz.) S.T. Blake] (2.1 ha), which originally was managed from 2004 to 2008 as part of the irrigated second ICL system described below but in 2009, was incorporated as part of the FRG CTN system to assist in cattle retention during drought years (Zilverberg, 2012). Steers grazed PNG, foxtail millet, and OWB1, before moving to the feedyard for finishing. Average stocking density for 2009–2010 in PNG, OWB1 and in the millet of the Mi–Ct rotation were 6.6, 14.5, and 5.7 steers ha−1, respectively. The PNG paddocks received a total of 60 kg ha−1 N, 11 kg ha−1 P and 18 kg ha−1 S in 2009, while no fertilizer was applied in 2010. In 2009 and 2010, OWB1 received an average of 200 mm of irrigation (via subsurface drip) and 477 mm of precipitation per year, which is equivalent to an average of 39% replacement of evapotranspiration (i.e., deficit irrigated). This paddock also received 67 kg N ha−1 and 12 kg S ha−1 each year in the last two years prior to sampling. Each crop of the millet–cotton rotation (Mi–Ct) received 56 kg N ha−1, 11 kg P ha−1 and 17 kg S ha−1 annually. The cotton variety was ‘FiberMax 9058F’.
- The second ICL (OWB_BER; 3.9 ha), initiated in 2003 and fully established (i.e., operational with grazing) in 2007, is a deficit-irrigated (average of 49% replacement of evapotranspiration) three-paddock agroecosystem consisting of two types of perennial warm-season grasses: ‘WW-B. Dahl’ old world bluestem and bermudagrass [Cynodon dactylon (L.) Pers.] (OWB BER). One paddock (2.1 ha) contains OWB (OWB2) and the remaining two paddocks contain ‘Tifton-85’ bermudagrass (BER) on 0.89 ha each. Because the two BER paddocks were similarly managed, only one bermudagrass paddock was sampled to conserve costs of analyses. Excess growth of bermudagrass was harvested for hay. All vegetative components of this agroecosystem and OWB1 from the FRG-CTN system were irrigated with a subsurface drip irrigation (SDI) system with tapes located on 1-m centers, buried 0.36 m deep. Irrigation in bermudagrass for 2009 and 2010 was 305 and 255 mm, respectively. Bluestem2 received 269 mm in 2009 and 175 mm in 2010. The OWB2 received 67 kg N ha−1 and 12 kg S ha−1, annually. The BER received 200 kg N ha−1 and 36 kg S ha−1 per year. During spring growth, OWB2 is grazed before BER. Average stocking density for 2009–2010 in OWB2 and BER were 21.8 and 67.4 steers ha−1, respectively. Grazing management and ICL system details are found in Zilverberg (2012).
- The third ICL (FRG_RC; 67.4 ha), 2.5 km northwest of Lockney, TX, consisted of rotational grazed (cow-calf) paddocks of bluestem (bluestem3 and bluestem3; 11.2 ha each) and 45 ha of fully irrigated annual crop production, which was corn (Zea mays; R-corn) in 2010 under conventional tillage and and fertilized through surface applications. Previous annual crops produced included cotton (2005 & 2007) and sunflower (2006 & 2009). Bluestem3 had been managed for grass forage since 2003 while bluestem4 was under annual crop production (cotton and corn followed by wheat planted for cover and grazing) until 2007 when bluestem was established for seed harvest and additional grazing. Total irrigation applied via center pivot (MESA) for the total agroecosystems was 178 and 228 mm in 2009 and 2010, respectively with the majority of the water applied to the crop area.
- The CTN1 agroecosystem (cotton1; 0.25 ha) was located on the Texas Tech Experimental Station, adjacent to the FRG_CTN and OWB_BER systems. It has been under conventionally tilled monoculture cotton since 2006 and was irrigated through SDI at 1 m spacing to a depth of 30 cm. The cotton variety was Delta Pine DP0912 B2RF, planted with Temik applied in furrows at 5.6 kg ha-1. In 2010 fertilizer (48.2 kg N ha-1) and irrigation (105 mm) were applied through the SDI system.
- The CTN2 agroecosystem (cotton2; 41.6 ha) located 1 km south of Lockney, TX used a reduced tillage practice with maximum tillage depth of 10 cm. Irrigation and fertilization were applied through SDI (1.5 m spacing buried to a depth of 25 cm) with more irrigation in 2009 (305 mm) compared to 2010 (127 mm).
Two additional agroecosystems (OWB_ROT and CTN) also located at the Texas Tech Experimental Farm, were assessed for aggregate stability and C dynamics (Fig 1B). The site was part of a non-irrigated grass-based grazing experiment (1986 to 1988) before being left unmanaged until 1997 when the two systems were initiated. Both systems were deficit irrigated using SDI with 1 m spacing at a depth of 0.36 m. The CTN3 agroecosystem consisted of continuous cotton with chemically terminated “Lockett” wheat (Triticum aestivum L.) planted in the furrows as a cover crop between cotton crops until 2004. The ICL (OWB_ROT) consisted of a cotton-forage-beef cattle system comprised of three vegetation components. Approximately half of the area (2.1 ha) contained a bluestem (bluestem5) paddock with the remaining area split between two paddocks (0.9 ha each) planted in a rotation of no-till cotton, wheat, fallow, and rye (Secale cereale L.) such that both paddocks did not have the same crop at the same time. Grazing steers were introduced in January of each year and rotated through the system until July when steers were moved to a feedlot.
Sample collection
In 1997, soil samples (0-15 cm) were collected from the additional ICL (OWB_ROT) and CTN systems (Fig 1B) prior to the initiation of the ICL and cotton plantin. These baseline samples were stored air-dried and subsampled for aggregate fractionation and C analysis in 2009. In January 2010 these fields were resampled (0-15 cm) by collecting a minimum of 4 subsamples per paddock which were gently composited to provide a representative sample at the vegetation level.
In July-August 2010, soil samples (0-5 and 5-20 cm) were collected from In the CTN1, CTN2, FRG_CTN, OWB_BER, and OWB_RC agroecosystems. Five soil samples were collected per rep/block using a zig-zag pattern, gently mixed by hand to avoid breaking aggregates and composited to provide a representative sample at the vegetation level. Soils were collected with a shovel from a shallow (22 cm) hole by removing a 5 cm thick by 13 cm wide slice of soil and trimming the edges to obtain a uniform amount of soil at each depth. This was method was used to minimize disruption to the soil aggregates.
All soils collected in 2010 were stored at 4°C and within 48 of collection, the field-moist soil was gently broken by hand to pass through an 8 mm sieve. Visible plant material was removed and a subsample was air-dried for 3 days for soil aggregate fractionation and chemical analysis. Another field-moist subsample was passed through a 4.75 mm sieve to be used for microbial community analysis.
A table is provided with an overview of the soil physical, chemical, and microbial assessments conducted (Table 1). Detailed information regarding each method is included in the text below.
Soil physical properties:
- Soil bulk density was measured at the CTN1, CTN2, FRG_CTN, OWB_BER, and OWB_RC agroecosystems using an InstroTek 3500 Xplorer Nuclear Moisture Density Gauge (Las Vegas, NV) at 5 and 20 cm. Bulk density was not measured on the OWB_ROT and CTN3 agroecosystems because 1997 data were not available.
- Particle size analysis was conducted by a combination of sieving and sedimentation techniques as described by Kettler et al (2001).
- Water-stable aggregate isolation: Four water stable aggregate fractions were obtained following a wet-sieving method of air-dried soils for all agroecosystems according to Elliott (1986). Prior to sieving samples were slaked by placing 100 g (250 µm) represented the small macroaggregates were transferred to a drying tin. The soil and solution which passed through the sieve was then poured onto a 53 µm sieve and the sieving process was repeated. Soil remaining the sieve (>53 µm) represented the microaggregate fraction, while the soil and solution which passed through the sieve represented the silt+clay fraction. The both the microaggregate and silt+clay fractions were transferred to drying tins. All aggregate fractions were dried at 60C and weighed to obtain the mass of each fraction.
- Mean weight diameter (an indicator of aggregate stability) was calculated based on the proportion of the soils with each of the water stable aggregate fractions. Calculations were done using the following equation:
MWD = Σi PiDi where Pi is the proportion of the whole soil in the given fraction, and Di is the average diameter (mm) of the particles of the fraction (van Bavel, 1949). - A modified method was used for field-moist soils to examine spatial distribution of microbial communities within soil aggregates. The fractionation method was followed as described above with the following modifications: all soils were maintained at field-moist conditions prior to sieving, all aggregate fractions were stored moist at 4C, ultrapure water with a DNAse/RNAsefree filter (Synergy UV, Millipore, MA) was used ruing sieving, sieves and containers were rinsed in a bleach solution between each sample, and the silt+clay fraction was collected using centrifugation (10 min at 5000 rpm and 4C) after addition of a weak (0.01M) CaCl2 solution.
Intra-aggregate isolation
- Three intra-aggregate fractions were isolated from the macroaggregate fraction according to Six et al (2000). Large and small macroaggregates were combined, subsampled, and placed in a device containing a 250 µm screen with 50 glass beads (4 mm). Through the combination of physical disturbance (shaking) and continuous water flow the macroaggregates were completely dispersed. Material passing through the screen was collected on a 53 µm sieve set above a catchment basing. Following dispersal the 53 µm sieve followed the wet-sieving process described above. Intra-aggregate particulate organic matter remained on the 250 µm screen, while intra-aggregate microaggregates remained on the 53 µm sieve and intra-aggregate silt+clay remained in the catchment basin.
- Because the CTN1, FRG_CTN, and OWB_BER were spatially separated from the CTN2 and OWB_RC fields sand correction was done for these agroecosystems. Sand correction was done by dispersing subsamples of the macroaggregate and microaggregate fractions in a 0.5% sodium hexametaphosphate solution while shaking for 15 hours.
Soil chemical analyses
- For the CTN1, CTN2, FRG_CTN, OWB_BER, and OWB_BER agroecosystems, chemical analyses was done through an independent lab (Ward Lab; Kearney, NE) and included pH (1:1) and soil organic matter (%LOI). Subsamples of each fraction and whole soil were hand ground to a fine powder and total nitrogen and total C content was determined by dry combustion analysis using a LECO TruSpec CN Analyzer (St. Joseph, MI). Presence of inorganic C (i.e. carbonate C) was examined through acidification with 1:1 HCl:H2O solution (Non-carbonate Carbon Method; LECO) of a subset of samples that covered the range of soil pH (6.9 to 8.3) prior to analysis. Following acidification, samples were analyzed using dry combustion and no detectable inorganic C was measured with the soil samples so that measured total C represents soil organic C. In the case where bulk density was measured it was used to determine total nitrogen soil organic C, and soil organic matter on an area basis (kg ha-1).
Greenhouse gas fluxes
- In the fall of 2009, 33 PVC collars (20.3 cm diameter by 11.4 cm tall) were placed with each replicate of PNG, Mi-Ct, Ct-Mi, BER, and OWB2. Three rings were placed in the PNG paddock and two rings were placed in the remaining paddocks. Each collar was buried to a depth of 8.9 cm and located in the center of each paddock and remained in place throughout the duration of the experiment with the exception of tillage and planting events in the cotton and millet rotations. Gas sampling occurred between 0800h and 1300h hours on all sampling dates with missing samples corresponding to management practices requiring the removal of collars (i.e. tillage or harvest).
Carbon dioxide measurements were collected using a LI-COR LI-8100a (Lincoln,NE) with a 20 cm survey chamber system interfaced with a Theta moisture (Dynamax; Houston, TX) and temperature probe. Gas measurements began in June of 2010 and continued every 7 days through September 2010) at which the sampling interval was increased to 30 days until September 2011. Due to the drought in 2011 all management practices were discontinued from May 2011 until 2012 although flux measurements continued. - Nitrous oxide fluxes were measured using a static chamber system (Hutchinson and Mosier, 1981) designed to allow for collection from the same collars used for CO2 flux measurements. On sampling dates, PVC caps (21.4 cm inner diameter) with two bulkhead fittings to allow for chamber venting and sample collection was placed on top of the collars. Chambers, at a height of 9.7 cm created a headspace of 2984 cm3 from which 30 mL samples were drawn at 15 minute intervals for 45 minutes. Gas samples were stored in syringes maintained under pressure and analyzed within 36 hours using a Shimadzu GC-2013 gas chromatograph (Kyoto, Japan). The GC was fitted with Restek packed column and an electron capture detector. Certified N2O standards (Air Liquide; Plumsteadville, PA) were used for calibration and net N2O fluxes were calculated as the linear change in concentration from time zero to 45 minutes. Additionally, soil temperature and volumetric soil moisture using a Campbell Scientific Hydrosense probe (Logan, UT) were collected at 10 cm depth for each sampling time.
Soil Microbial Analyses
- Microbial biomass C and N: Soil microbial biomass C (MBC) and N (MBN) were assessed via the chloroform-fumigation extraction method (Brookes et al., 1985; Vance et al., 1987) using 15 g of oven-dry equivalent field-moist samples.
- Microbial community composition using EL-FAME profiles: Field-moist soil samples were analyzed according to the ester-linked fatty acid methyl esters (EL-FAMEs) method by (Schutter and Dick, 2000). The FAME fractions were dried under N2 and analyzed using a 6890 GC series II (Hewlett Packard, Wilmington, DE, USA) equipped with a flame ionization detector and a fused silica capillary column (25 m × 0.2 mm) and H2 (ultrahigh purity) as the carrier gas (Acosta-Martínez et al., 2010). Peak identification and area calculation was performed using the TSBA6 aerobe program from MIDI (Microbial ID, Inc., Newark, DE). Absolute amounts of FAMEs (nmol g−1 soil) were calculated according to (Zelles, 1997) Zelles (1997) using the 19:0 internal standard and these values were subsequently used to calculate mol percent. The FAMEs are described by the number of C atoms, followed by a colon, the number of double bonds and then by the position of the first double bond from the methyl (ω) end of the molecule. Arbuscular mycorrhizal fungi (AMF) were categorized by the FAME markers 16:1 ω 5c and 20:1 ω9c (Graham et al., 1995; Madan et al., 2002; Olsson, 1999) and saprophytic fungi using the FAME makers 18:1ω9c, 18:2ω6c and 18:3ω6c (Frostegård and Bååth, 1996; Zelles, 1997; Zelles et al., 1991). Because the 18:2ω6,9 and 18:1ω9 markers for saprophytic fungi are also major plant fatty acids (Frostegård et al., 2011), care was taken when processing samples for FAME analysis to remove all visible plant parts to minimize this interference.
- Soil saprophytic fungal functional diversity using C utilization profiles: Fungal C utilization profiles were assessed via the FungiLog procedure, as described by (Dobranic and Zak, 1999) Dobranic and Zak (1999), which uses Biolog SFN2 96-well microtiter plates containing 95 different C substrates (Biolog, Hayward, CA, USA). Each well was inoculated with a 100-µl mixture of water, agar, streptomycin sulfate and chlortetracycline hydrochloride (used as antibiotics), dimethylthiazolyl-diphenyltetrazolium bromide (color reaction dye) and 50 mg of soil OM particles between 250 and 500 µm in size (Sobek and Zak, 2003). The microtiter plates were incubated at room temperature for 120- h (time which allows for the most complete C utilization) and then absorbance was measured at 590 nm on an Epoch spectrophotometer (BioTek, VT, USA). Absorbance values were converted to fungal functional responses for total substrate activity (SA), substrate richness (SR) and substrate diversity (H) as described by (Zak et al., 1994) Zak et al. (1994). Briefly, SA is calculated as the sum of the absorbance across the entire plate and represents the combined capacity to utilize different C substrates; SR is the total number of wells displaying any activity; and H is equivalent to the Shannon index (Magurran, 1988) and is calculated as the ratio of the activity (absorbance at 590 nm) of a particular C substrate to the sum of all absorbance values > 0 (Dobranic and Zak, 1999). Each C substrate on the plate was analyzed individually and collectively as one of six groups (guilds) of chemically similar substrates. The six guilds were: (1) amines/amides, (2) complex carbohydrates, (3) carboxylic acids, (4) polymers, (5) amino acids and (6) simple carbohydrates (Sobek and Zak, 2003).
- Enzyme assays (for nutrient cycling assessment): Soil enzymatic activities involved in C-cycling (β-glucosidase, α-galactosidase, and β-glucosaminidase), P-cycling (alkaline phosphatase and phosphodiesterase) and S-cycling (arylsulfatase) were used as an assessment of soil metabolic functionality. The enzymatic activities were assayed using 0.5g of air-dried soil under final concentration of 10mM of the specific enzyme substrate (p-nitrophenyl derivate) and buffer, and incubated (37?C) at their optimal pH as described in Tabatabai (Tabatabai, 1994) and Parham and Deng (2000). The concentration of the reaction product was determined colorimetrically at 400nm using an Epoch spectrophotometer (BioTek Instruments, VT, USA). The results for enzyme activities were expressed in mg of p-nitrophenol (PN) released per kg−1 soil h−1. All enzyme activities were assayed in duplicate with one control, to which substrate was added after incubation and subtracted from a sample value.
- DNA extraction and Pyrosequencing: DNA was extracted from 0.7 g of moist soil using the Fast DNA Spin Kit for soil (MP Biomedicals, OH, USA) according to the manufacturer’s instructions. The extracted DNA (1µl) was quantified using an Epoch plate spectrophotometer (BioTek Instruments, VT, USA). The ratio of the absorbance at 260 and 280 nm (A260/280) was used to assess the purity of nucleic acids with acceptable values considered at or above ~1.8. All DNA samples were diluted to 30ng/μl for a 50 µl PCR reaction.
Chimera detection and removal was performed using specialized pipeline executing UCHIIME in de novo mode on the clustered data that was output of denoising methods. Sequences that pass the quality control screening are condensed into a single FASTA formatted sequence. These sequences are then clustered into OTU clusters with 100% identity (0% divergence) using USEARCH (Edgar, 2010). For each cluster the seed sequence was placed into a FASTA formatted sequence file and queried against a database of high quality sequences derived from NCBI using a distributed .NET algorithm that utilizes BLASTN+ (KrakenBLAST www.krakenblast.com). Using a .NET and C# analysis pipeline the resulting BLASTN+ outputs were compiled and data reduction analysis performed as described previously (Andreotti et al., 2011; Bailey et al., 2010; Callaway et al., 2010).
Experimental design
Four of the agroecosystems (FRG_CTN, OWB_BER, CTN3, and OWB_ROT) incorporated true replication via establishment of 3 true blocks in a randomized block design. The remaining three agroecosystems (CTN1, CTN2, and OWB_RC) had no true replication in spaces, and thus 3 pseudo-replicates were established within the area of the agroecosystems.
{kind=link}
Results and discussion of key findings are organized accorging to our three objectives.
1) Measure the amount of C stored in active, intermediate and passive SOM pools using a detailed physical fractionation method, which provides up to eight different soil aggregate fractions.
The main goal of this objective was to identify agroecosystems that had the greatest potential for C sequestration. Soil organic matter is comprised of various pools with differing physical and chemical properties that turnover and are cycled through the system at different time scales. Organic C can be physically and chemically protected within different sized aggregates and thus, it is necessary to evaluate soil organic C distribution within and among aggregate fractions. In this study, six soil aggregate C pools were physically isolated to evaluate the C sequestration potential of three ICLs and two continuous cotton (CTN) agroecosystems in the Texas High Plains. The three ICLs investigated were the FRG-CTN, OWB-BER, and OWB-RC systems. Among the fractions, the macroaggregates are of particular interest because they function to resist the forces of water and wind and therefore reduce erosion risks. The macroaggregates also serve as a protective shell for SOC in smaller microaggregates found within the macroaggregates. Carbon stored within these intra-aggregate microaggregates (intra-aggregate microaggregate) have been proposed as an indicator of C sequestration. Thus, macroaggregates serve two important ecological functions: resistance to erosive forces and C sequestration potential. Another measure of aggregate stability is mean weight diameter, which is calculated based on the proportions of the macroaggregates, microaggregates, and silt+clay fractions. The results for this objective have been published recently (Fultz et al., 2013b) and are summarized in the following three points:
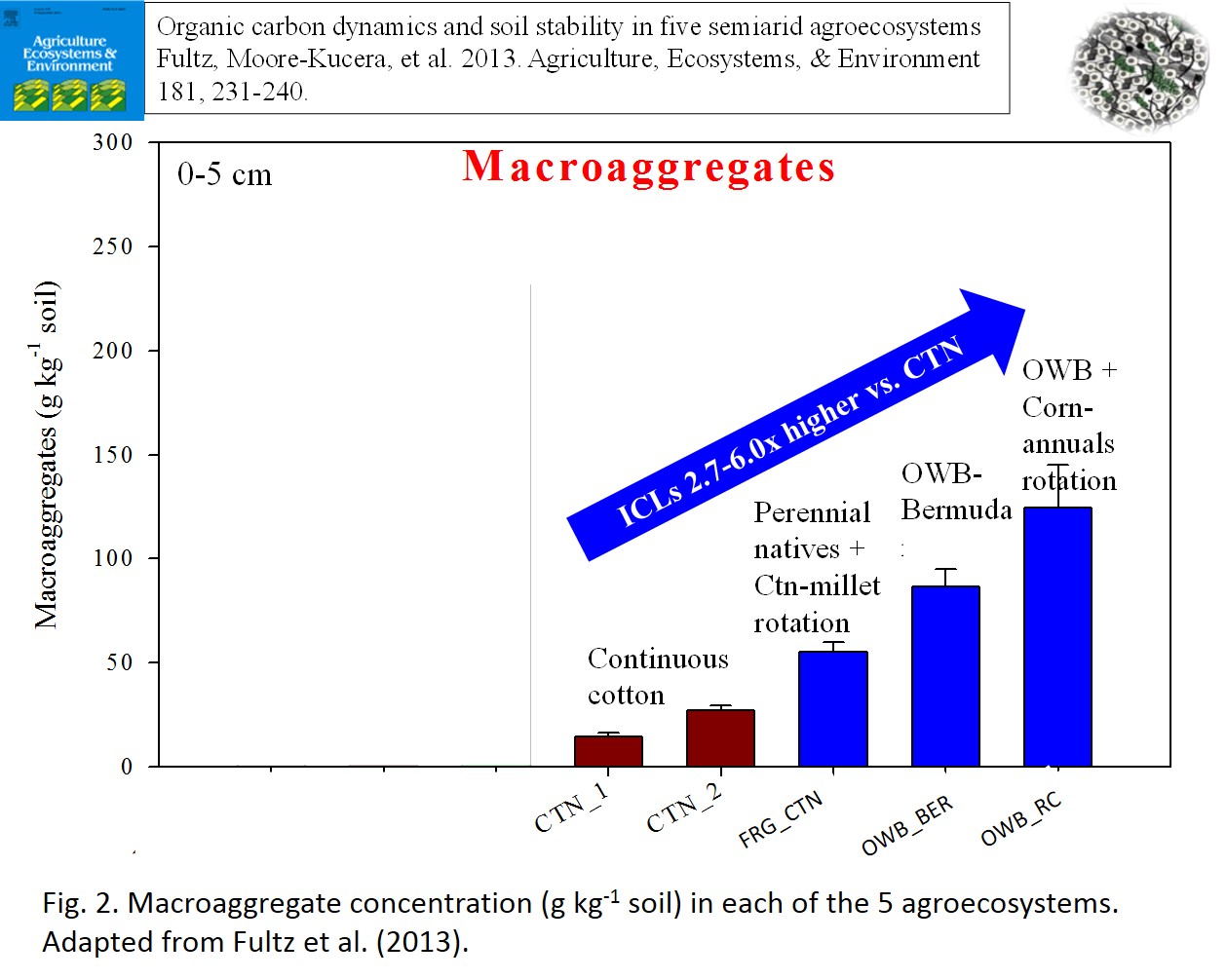
- Relative to the continuous cotton systems, the ICL agroecosystems increased the proportion of the water stable macroaggregates (Fig 2), and mean weight diameter, which provided a physical protective shell for SOC via the formation of intra-aggregates within the macroaggregates. However, there was a gradient in macroaggregate concentration across the ICL systems, which tended to align with irrigation practices (Fig 2). The ICLs under dryland had the lowest amount of macroaggregates, MWD, and C stored in the protected intraggregate microaggregates. The deficit irrigated ICL was intermediary and the producer-operated FRG_RC ICL that had half of the land under deficit-irrigated OWB and the other half fully irrigated annual rotations had the greatest levels. These results indicate that there was great flexibility in achieving both productive crops and cattle outputs as well as ecological benefits; thus giving producers options to optimize wide-scale system goals.
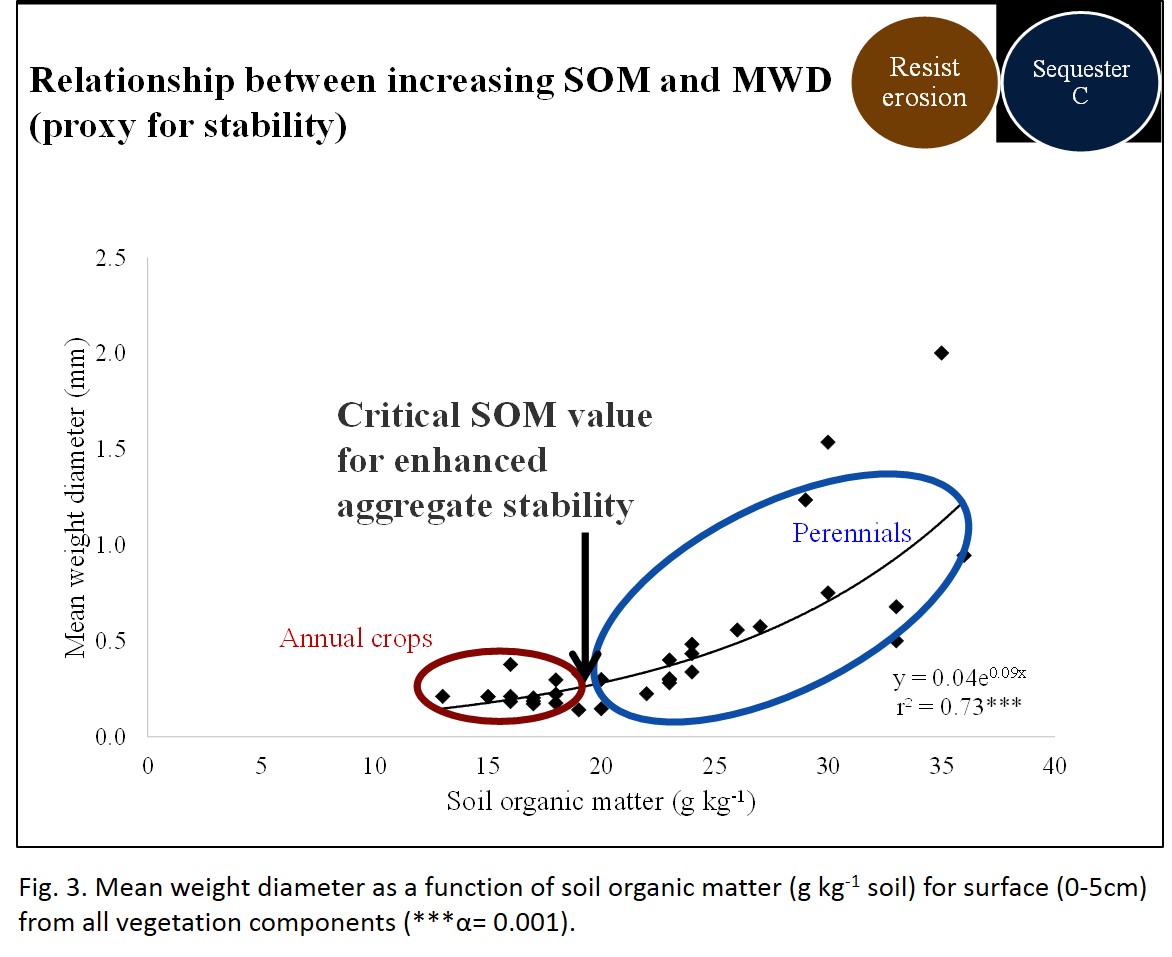
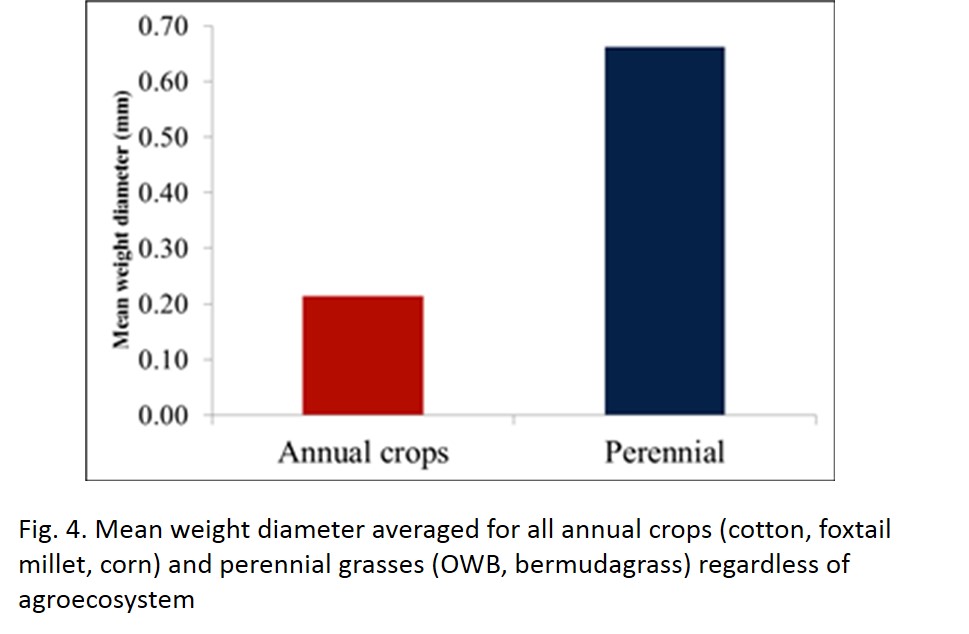
- Systems that could sustain a SOM level of above 1.7% (equivalent to 17 g kg-1) or a SOC level of at least 5.5 Mg ha-1, were strongly correlated with enhanced mean weight diameter (MWD), and thus aggregate stability (Fig. 3). The values below this threshold were primarily associated with the annual crops (cotton, foxtail millet, corn, etc.) and those above the threshold were associated with fields under perennial grasses (e.g., OWB, bermudagrass, perennial native grasses). Fig 4 depicts the average MWD for annual and perennial vegetation components regardless of agroecosystem. Although specific to these semi-arid soils, this critical level can act as a target for producers to minimize and ultimately reverse soil degradation.
- The vegetation components of the agroecosystems with the greatest C sequestration potential (as measured by intra-aggregate C) and soil stability (as measured by MWD) included perennial grasses and irrigated diverse crop rotations (R-corn). This is especially important in the Southern High Plains regions because it implies that as water resources decline, producers have options to assist during the transition from 100% conventional, fully irrigated monoculture cotton to dryland or deficit-irrigated crops or ICLs. These benefits impart important ecosystem services such as C storage and reduced erosion potential and complement the benefits previously reported regarding energy, water, and nutrient conservation (Fultz et al., 2013b).
C dynamics summary from the additional sampling (OWB_ROT and CTN):
A continued increase in SOC was measured in the ICL following 13 yr of management with 22% more SOC relative to CTN. In contrast, SOC declined almost 3 times the original levels when converted to continuous cotton management. The ICL under limited irrigation enhanced SOC stored in protected, recalcitrant aggregate pools (intra-aggregate microaggregate SOC of 8.2 and 5.4 mg g-1 macroaggregate in the ICL and CTN, respectively). These benefits impart important ecosystem services such as potential C sequestration and reduced erosion potential, which are especially important in these semiarid soils.
2) Evaluate greenhouse gas fluxes (CO2 and N2O) to calculate global warming potential within each system.
The ICLs described in objective 1 results above supported increased whole SOC and C protected within aggregates inside of the macroaggregate shells. Offsets to enhanced C-sequestration however can occur via carbon dioxide (CO2) and nitrous oxide (N2O) emissions. The purpose of this study was to examine the magnitude and seasonal patterns of CO2 and N2O fluxes associated with the FRG_CTN and OWB_BER ICLs.
Measured N2O fluxes were episodic in nature and sensitive to increased moisture, SOC content, and fertilization rates. N2O fluxes ranged from -17 to 113 µg N2O-N m-2 h-1 with only three significant increases in flux measured all occurring in the BER paddock. The greater MWD measured in the BER component is indicative of greater proportions of large aggregates and supports the proposed relationship between aggregation, porosity, and increased fluxes. Greater fluxes in no-till systems have been attributed to lower gas diffusion rates from the soils due to increased water-filled pore space and greater quantities of larger aggregates (Ball et al., 1999). While denitrification is typically considered the process most responsible for measured emissions of N2O, in these semi-arid systems nitrification by heterotrophic organisms are likely a major driving factor. Through the use of subsurface irrigation and lower precipitation within the Southern High Plains region, saturation at the soil surface is a rare occurrence. For this reason, denitrification is most likely to occur within anaerobic microsites at depth where oxygen may become limiting. Additionally, in the case of the D_OWB and BER, nitrogen is not likely to be limiting due to the addition of N-fertilizer. However, application of readily available N through subsurface drip irrigation ensures decreased availability within surface soils and decreases or delays the expected contribution through nitrification. Significant spikes in the BER component occurred within two weeks of fertigation events, which may contribute to the release of N2O via the nitrification process. The existence of negative flux rates would indicate that there may be times at which atmospheric N2O is being taken up by soil organisms resulting in a potential sink (Rafique et al., 2011). Within these systems, N2O production did not contribute significantly to the global warming potential as the highest measured N2O emissions corresponded to CO2 flux rates measured during the period of lowest activity. The global warming potential for N2O converted to CO2 equivalents ranged from -45 to 306 g CO2-C equivalents m-2 yr-1. In contrast, CO2 GWP ranged from 103 to 4451 g CO2-C m-2 yr-1 over the time period of measurement. Therefore, N2O does not appear to significantly contribute to the GWP in these semi-arid soils.
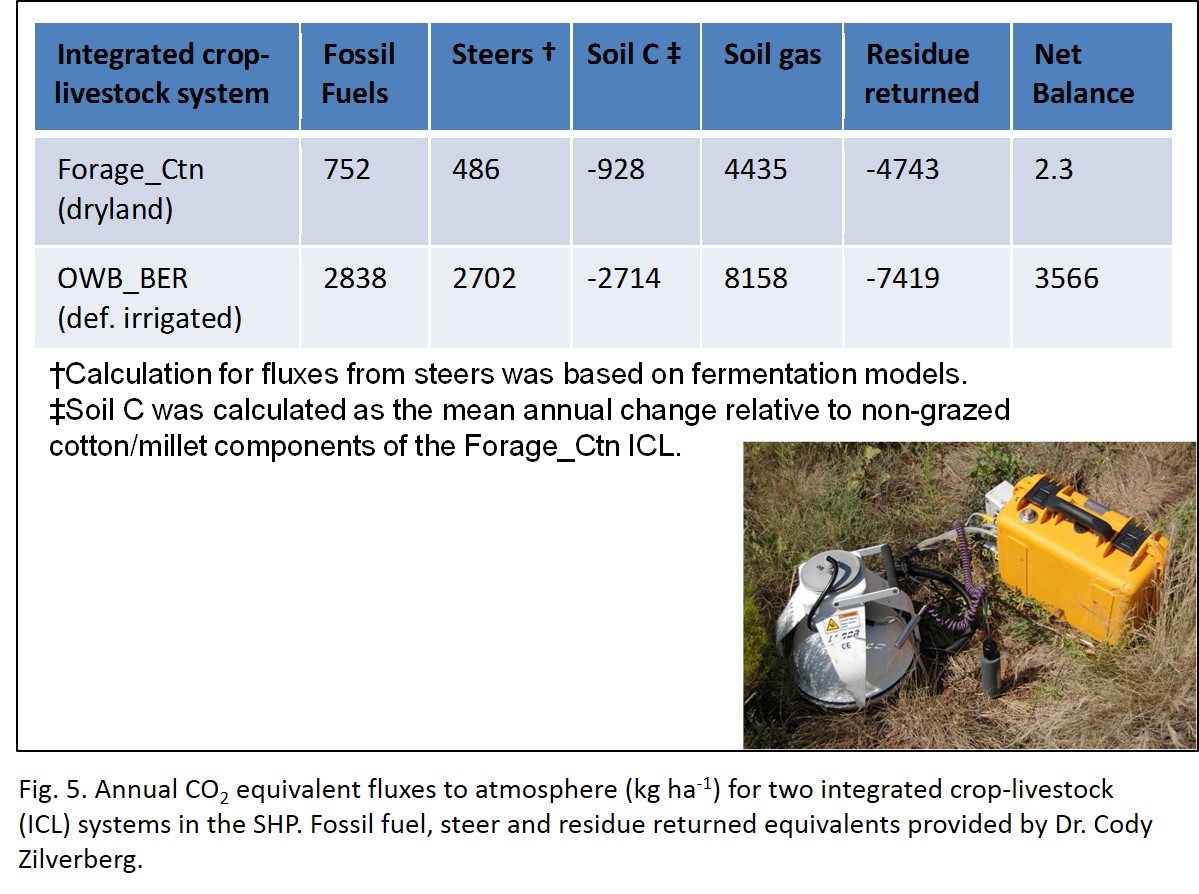
Working in collaboration with another PhD student (Dr. Cody Zilverberg) who was under the guidance of Dr. Vivien Allen, we calculated the annual CO2 equivalent fluxes to the atmosphere (kg ha-1) for the FRG_CTN and OWB_BER ICLs (Fig. 5). Although the OWB_BER was characterized by greater storage of soil C, the FRG_CTN had essentially no net release of CO2-Ceq (2.3 kg ha-1) compared to 3566 kg ha-1 CO2-Ceq released to the atmosphere by the OWB_BER system. The greater flux in this system was mainly attributed to the high fossil fuel consumption (primarily from N fertilization). This suggests that more research is needed to identify the most efficient and lowest inputs of N to support this system if a lower CO2-Ce output is desired.
3) Characterize soil microbial community structure using fatty acid methyl ester (FAME) profiling and microbial biomass C (MBC) including bacterial diversity in whole soil and different soil aggregate fractions using a novel molecular biological tool (i.e., pyrosequencing).
Differences in the community structure, metabolic functioning (i.e., nutrient cycling potential via enzyme activities) among the agroecosystems can be summarized in the following points.
MBC, FAME, and Enzyme Summary
The ICL agroecosystems (OWB_BER and FRG_CTN) supported increased MBC (mg kg-1 soil), enzyme activities involved in C, N, P, and S cycling, total FAME concentration (nmol g-1 soil), and specifically higher absolute (nmol g-1) and relative abundance (mol %) of arbuscular mycorrhizal fungi (AMF) and saprophytic fungi compared to continuous cotton (CTN). Ranking among the three systems for these properties followed the order OWB_BER > FRG_CTN > CTN.
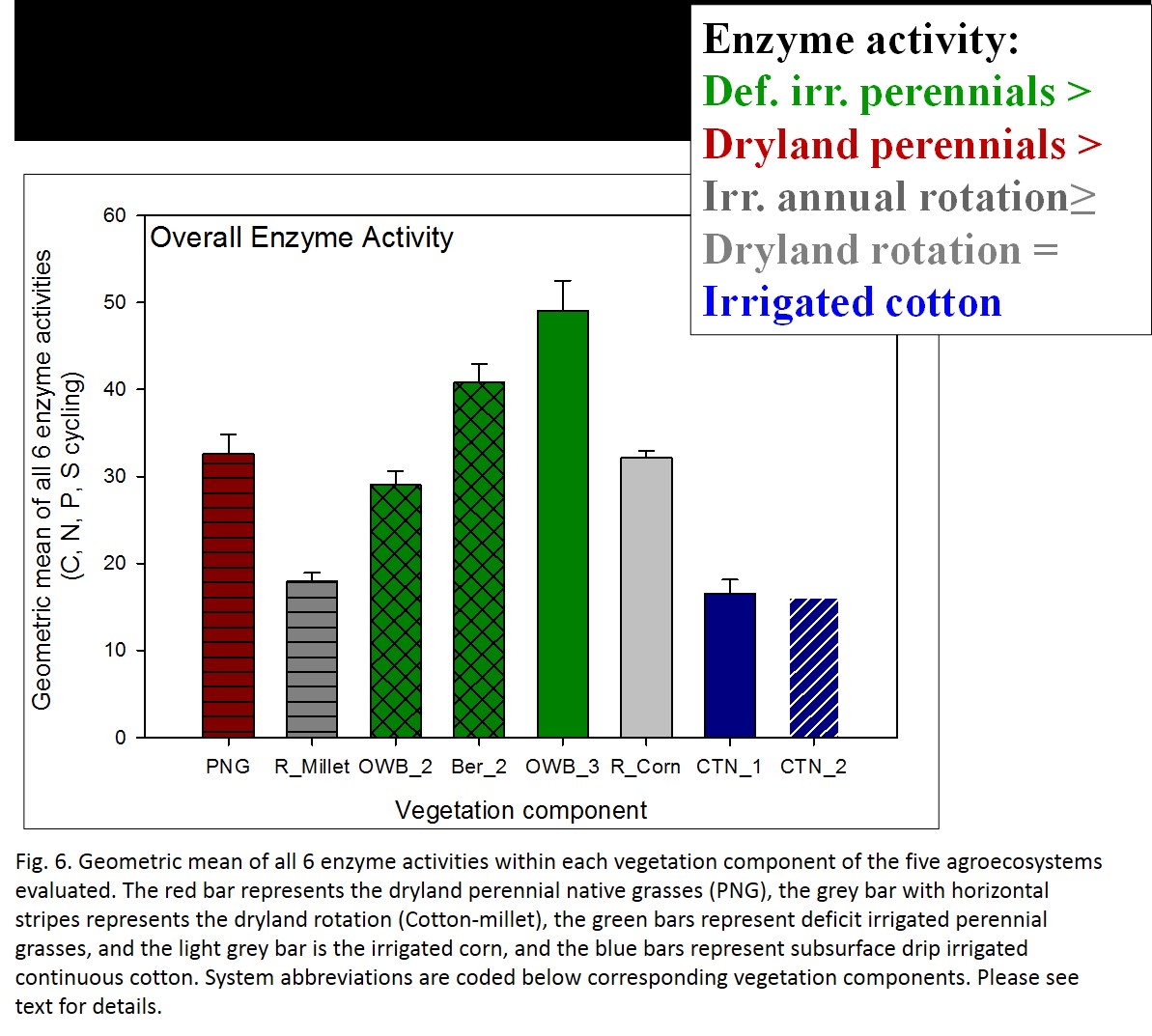
The geometric mean of all enzyme activities (6 enzyme activities involved in C, N, P, and S cycling) was greatest in the OWB_BER system and lowest in the continuous cotton systems (Fig. 6). At the vegetation level, perennials had higher activity, followed by crops under rotation, and monoculture cotton had the lowest level geometric enzyme mean.
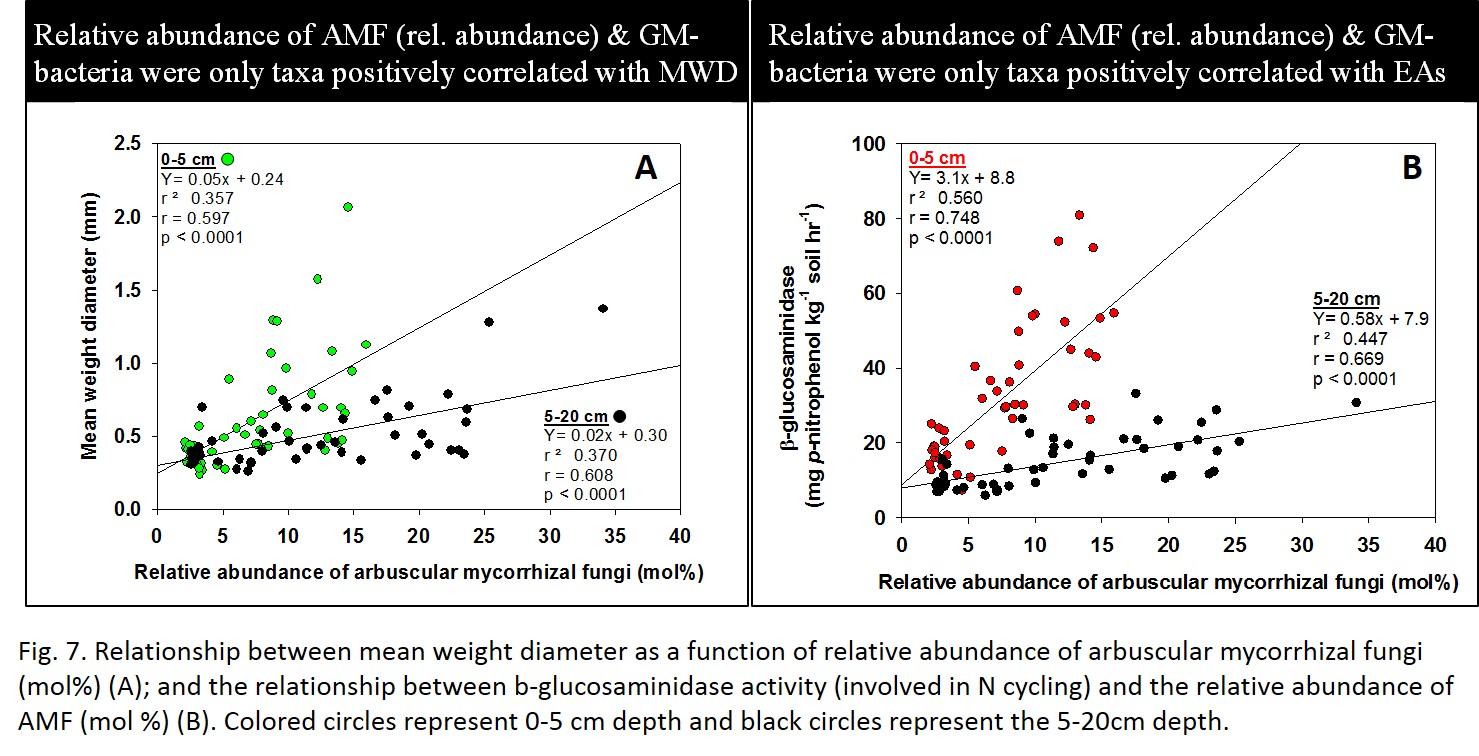
The absolute abundance (nmol g-1 soil) of all FAME microbial groups (e.g., Gram positive and negative bacteria, total bacteria, AMF, saprophytic fungi, etc) were positively correlated with increased mean weight diameter (MWD; a proxy for aggregate stability). This finding was not surprising because microorganisms are key agents in creating stable aggregates (i.e., as microbial biomass increases, MWD should also increase). However, what was of ecological significance was the positive correlation between the relative abundance (mol %) of AMF and Gram negative bacteria and MWD (Fig 7A). This also was true for all enzyme activities (e.g., Fig 7B). In contrast, the relative abundance of all other FAME groups were negatively correlated. These findings are of ecological significance because they provide evidence to link specific microbial taxa (AMF and Gram negative bacteria) to ecosystem function (i.e., enhanced soil stability and nutrient cycling. Therefore, tracking of these microbial taxa may be serve as sensitive indicators for agroecosystem functioning.
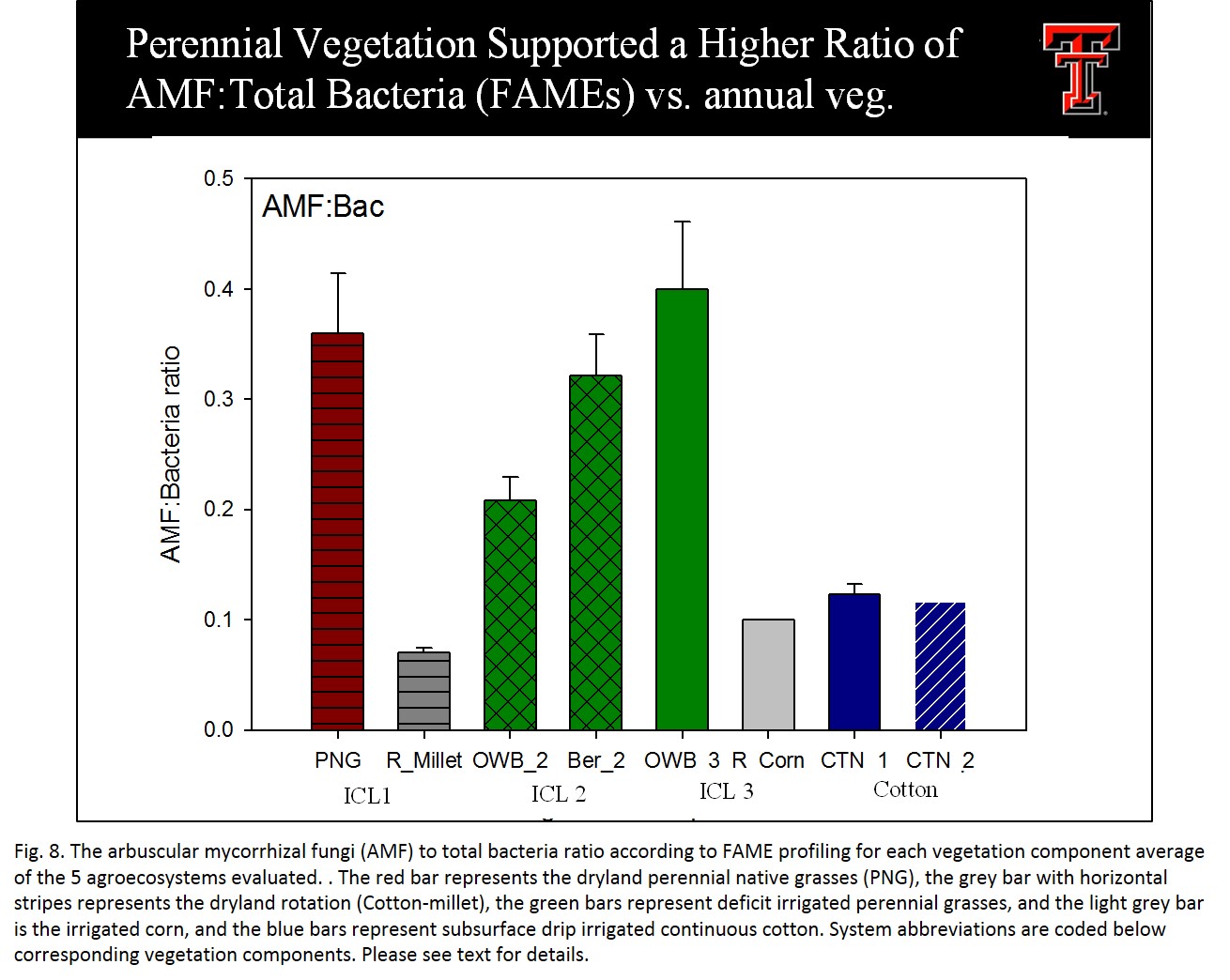
Among vegetation components, perennial vegetation supported a higher ratio of AMF:Total Bacteria (measured by FAME profiling) compared to the annual crops (cotton, millet, corn). This ratio may provide a sensitive indicator of agricultural systems that lead to enhanced aggregate stability and nutrient cycling (Fig. 8).
Fungal and Bacterial Diversity Summary
Fungal pyrosequencing identified on average 2484 OTUs per sample, out of these 87% were Ascomycota, 6% were Basidiomycota, and Glomeromycota was 1%. At the order level, the FRG_CTN system was characterized by the highest abundance of Hypocreales, Mortierellales dominated OWB_BER, while OWB_RC had higher Glomerales compared to other systems. Filobasidiales dominated CTNc’s, while Mi-Ct had highest abundance of Hypocreales.
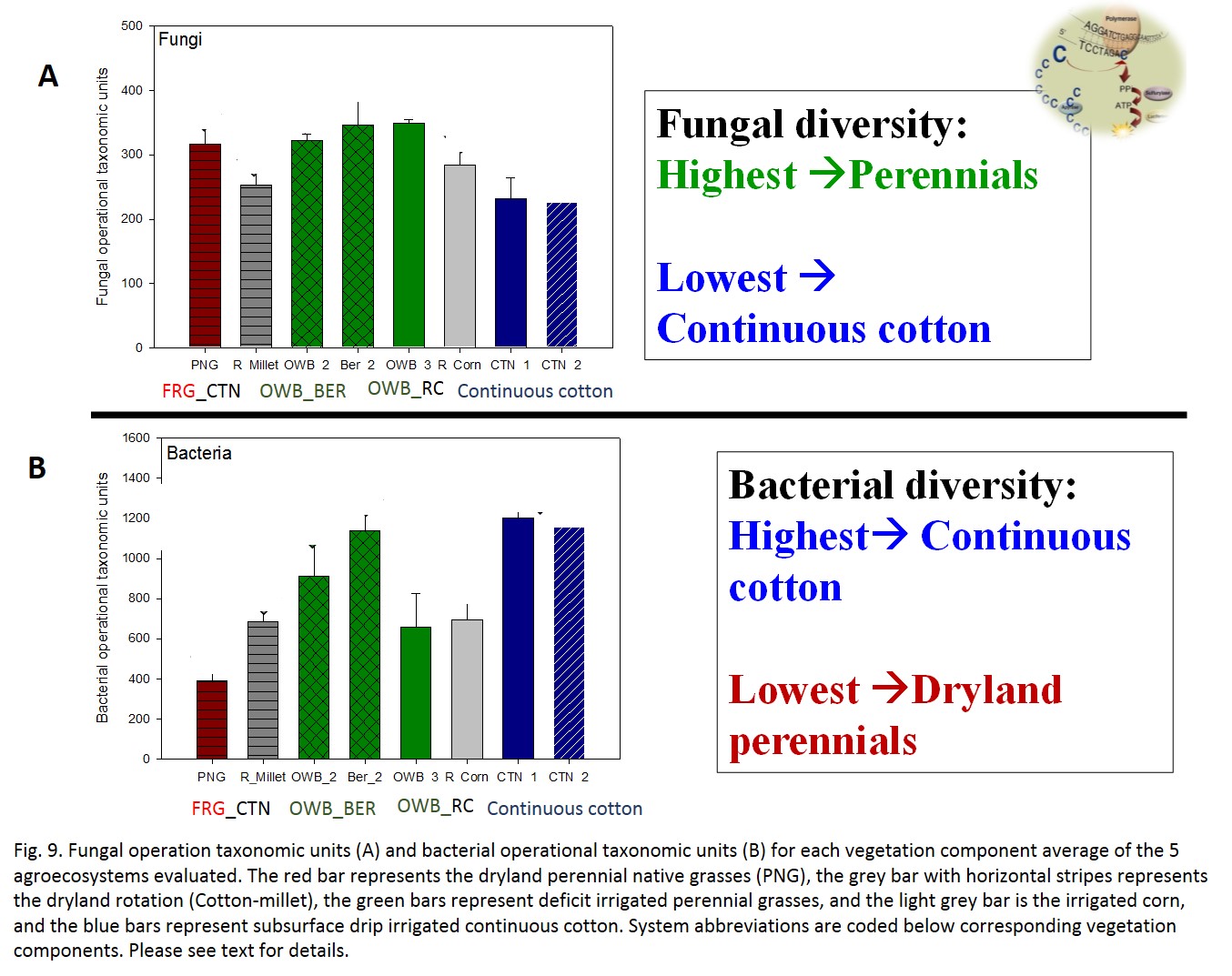
Fungal diversity indexes were sensitive to agroecosystem and vegetation components (Fig. 9A). In general, the two continuous cotton systems had lower fungal diversity measures than the ICLs systems, although the differences were not always significant at alpha = 0.05. OWB_BER had the highest operational taxonomic units (OTUs) (Fig 9A). Fungal diversity (OTUs) was higher under OWB_BER and lower under CTN_2, while bacterial diversity was higher under CTN_1, 2 and lower under FRG_CTN and OWB_RC (Fig 9B). These divergences between fungi and bacteria diversity measures suggest broad functional differences in the contribution of these microbial groups to ecosystem functioning. For example, fungi may play a more important role in the ICL systems, especially non irrigated or deficit irrigated vegetation because of the role of fungi in transporting and redistributing water within the soil profile. Additionally, bacteria (especially, the Actinobacteria and other Gram positive bacteria are important in low organic matter soils for the degradation of more recalcitrant compounds.
Bacterial pyrosequencing identified on average 1733 OTUs per sample, representing Proteobacteria (35%), Actinobacteria (32%) and to lesser extent Bacteroidetes (7%) and Chloroflexi (8%). Specific bacterial taxa distinguished the agroecosystems from each other. For example FRG_CTN had the highest abundance of Actinobacteria, OWB_BER system was dominated by α, β-Proteobacteria and Chloroflexi. The continuous cotton systems were dominated by Bacteroidetes, while Mi-Ct rotation had high Actinobacteria.
One measure of the relative contribution of fungi and bacteria, is the ratio of the fungal OTUs to bacterial OTUs (F:B). This comparison revealed that the dryland perennial native grass paddocks had the highest F:B ratio and the continuous cotton fields had the lowest F:B ratio (data not shown). Fungi are well known for symbiotic associations with plants to assist with water and nutrient uptake. This may explain the greater ratios found in the dryland perennial fields compared to the irrigated cotton fields. These associations may play vital roles in the resilience and resistance of semi-arid ecosystems to drought and low phosphorus availability (because of the high alkaline conditions found in these soils). In fact, there was a strong positive relationship between the relative abundance (mol%) of arbusuclar mycorrhizal fungi (as measured by FAME profiling) and all enzyme activities including the three phosphorus cycling enzymes.
- Fig 3. Mean weight diameter (measure of aggregate stability) as a function of soil organic matter.
- Fig 4. Mean weight diameter averaged for all annual crops (cotton, foxtail millet, corn) and perennial grasses (OWB, bermudagrass) regardless of agroecosystem
- Fig 9. Fungal and Bacterial Operational Taxonomic Units (measure of diversity) in each vegetation component of the five agroecosystems evaluated.
- Fig 2. Macroaggregate concentration (g kg-1 soil) in each of the 5 agroecosystems.
- Fig 5. Estimate of global warming potential for two integrated crop-livestock systems (FRG_CTN andOWB_BER)
- Fig 7. AMF vs MWD and AMF vs beta-glucosaminisase activity
- Fig 8. The arbuscular mycorrhizal fungi (AMF) to total bacteria ratio according to FAME profiling for each vegetation component average of the 5 agroecosystems evaluated.
- Fig 6. Geometric mean of all 6 enzyme activities within each vegetation component of the five agroecosystems evaluated.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Educational & Outreach Activities
Participation summary:
Publications
- Davinic, M., Fultz, L.M., Acosta-Martinez, V., Calderon, F., Cox, S., Dowd, S., Allen, V., Zak, J., and Moore-Kucera, J. 2012. Pyrosequencing and mid-infrared spectroscopy techniques reveal distinct aggregate stratification of soil bacterial communities and organic matter composition. Soil Biology and Biochemistry 46:63-72. doi:10.1016/j.soilbio.2011.11.012. http://www.sciencedirect.com/science/article/pii/S0038071711004044.
- Davinic, M., J. Moore-Kucera, J., Acosta-Martinez, V., Zak, J., and Allen, V. 2013. Soil fungal distribution and functionality as affected by grazing and vegetation components of integrated crop-livestock agroecosystems. Applied Soil Ecology 66:61-70. doi:10.1016/j.apsoil.2013.01.013. http://www.sciencedirect.com/science/article/pii/S0929139313000401.
- Fultz, L.M., Moore-Kucera, J., Zobeck, T.M., Acosta-Martinez, V., and Allen, V.G. 2013. Aggregate carbon pools after 13-years of integrated crop-livestock management in semi-arid soils. Soil Science Society of America Journal 77:1659-1666. doi: 10.2136/sssaj2012.0423. https://www.soils.org/publications/sssaj/abstracts/77/5/1659.
- Fultz, L.M., Moore-Kucera, J., Zobeck, T.M., Acosta-Martinez, V., Wester, D.B., and Allen, V.G. 2013. Organic carbon dynamics and soil stability in five semiarid agroecosystems. Agriculture, Ecosystems, and Environment 181:231-240. doi: 10.1016/j.agee.2013.10.004. http://www.sciencedirect.com/science/article/pii/S0167880913003423.
Dissertations
- Fultz, L.M. August 2012 Dynamics of soil aggregation, organic carbon pools, and greenhouse gases in integrated crop-livestock agroecosystems in the Texas High Plains. Texas Tech University. http://hdl.handle.net/2346/48715.
- Davinic, M. December 2012. Soil microbial community diversity and functionality as affected by integrated cropping-livestock systems in the Southern High Plains. http://hdl.handle.net/2346/50744.
Conferences/Abstracts/Proceedings:
- Moore-Kucera, J., Davinic, M.*, Fultz, L.M.*, Acosta-Martinez, V., Dowd, S.E., Cox, S., Allen, V., 2010. Pyrosequencing-based assessment of bacterial diversity in soil aggregates under irrigated crop and livestock systems. The 2nd Annual Argonne Soils Workshop (Oct 6-8), Argonne, IL.
- Davinic, M.*, Moore-Kucera, J., Fultz, L.*, Dowd, S., Acosta-Martinez, V., Cox, S.B., Allen, V., 2011. Pyrosequencing-based assessment of soil bacterial communities within soil aggregates: Linking structure to C storage. Abstracts, Ecological Society of America Annual Meeting (August 11), Austin, TX. http://eco.confex.com/eco/2011/webprogram/Paper31444.html
- Fultz, L.M.*, Davinic, M.*, Allen, V., Moore-Kucera, J., 2011. Long-Term integrated crop-livestock agroecosystems and the effect on soil carbon. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 19), San Antonio, TX. http://a-c-s.confex.com/crops/2011am/webprogram/Paper65233.html
- Fultz, L.M.*, Davinic, M.*, Allen, V., Moore-Kucera, J., 2011. Dynamics of soil aggregation and carbon in long-term integrated crop-livestock agroecoystems in the Southern High Plains. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 17), San Antonio, TX. http://a-c-s.confex.com/crops/2011am/webprogram/Paper65237.html
- Fultz, L.M.*, Davinic, M.*, Cornejo, F.*, Allen, V., Moore-Kucera, J., 2011. CO2 and N2O fluxes in integrated crop livestock systems. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 17), San Antonio, TX. http://a-c-s.confex.com/crops/2011am/webprogram/Paper65081.html
- Davinic, M.*, Fultz, L.M.*, Acosta-Martinez, V., Calderon, F., Allen, V., Dowd, S.E., Moore-Kucera, J., 2011. Aggregate stratification assessment of soil bacterial communities and organic matter composition: Coupling pyrosequencing and mid-infrared spectroscopy techniques. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 18), San Antonio, TX. http://a-c-s.confex.com/crops/2011am/webprogram/Paper66342.html
- Davinic, M.*, Fultz, L.M.*, Acosta-Martinez, V., Zak, J., Allen, V., Moore-Kucera, J., 2011. Soil fungal community and functional diversity assessments of agroecosystems in the Southern High Plains. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 17), San Antonio, TX. http://a-c-s.confex.com/crops/2011am/webprogram/Paper66392.html
- Davinic, M.*, Fultz, L.M.*, Acosta-Martínez, V., Allen, V., Dowd, S.E. and Moore-Kucera, J. 2011. Soil microbial dynamics in alternative cropping systems to monoculture cotton in the Southern High Plains. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 17), San Antonio, TX. http://a-c-s.confex.com/crops/2011am/webprogram/Paper66308.html
- Fultz, L.M.*, Allen, V., Moore-Kucera, J., 2011. Long-term integrated crop-livestock agroecosystems in the semi-arid Southern High Plains and their influence on soil carbon. Abstracts, Annual Meeting of the American Geophysical Union (December), San Francisco, CA. http://adsabs.harvard.edu/abs/2011AGUFM.B31F0372F
- Zilverberg, C.J.**, Fultz, L.M.*, Michael, G., Moore-Kucera, J., Allen, V.G., 2012. Carbon release from steers and fossil fuels in integrated crop-livestock systems. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 23), Cincinnati, OH. http://scisoc.confex.com/scisoc/2012am/webprogram/Paper71018.html
- Fultz, L.M.*, Moore-Kucera, J., Allen, V., 2012. Significant management impacts on intra-aggregate soil fractions. Abstracts, ASA, CSSA, & SSSA International Annual Meetings, (October 23), Cincinnati, OH. http://scisoc.confex.com/scisoc/2012am/webprogram/Paper71011.html
- Davinic, M.*, Acosta-Martínez, V., Allen, V., Zak, J., Moore-Kucera, J., 2012. Soil fungal dynamics in perennial and annual crops of integrated crop and livestock systems. Abstracts, ASA, CSSA, & SSSA International Annual Meeting (October 23), Cincinnati, OH. http://scisoc.confex.com/scisoc/2012am/webprogram/Paper71013.html
- Calderon, F.J., Fultz, L.M.*, Allen, V., Moore-Kucera, J. 2012. Soil carbon and soil organic matter quality in soil size fractions from crop and livestock systems in Texas. Abstracts, ASA, CSSA, & SSSA International Annual Meetings, (October 23), Cincinnati, OH. http://scisoc.confex.com/scisoc/2012am/webprogram/Paper71012.html
- Fultz, L.M.*, Moore-Kucera, J., Allen, V., 2012. Increases in protected soil organic carbon found in perennial grassland vegetation as part of integrated crop-livestock systems. Annual Meeting of the Ecological Society of America (August 9), Portland, OR.
- Davinic, M.*, Fultz, L.M.*, Moore-Kucera, J., Acosta-Martinez, V., Zak, J., Allen, V., 2013. Soil fungal community and functional diversity in southern high plains integrated crop and livestock systems. Soil Ecology Society Conference (June 11-14), Rutgers University, Camden, NJ.
- Moore-Kucera, J., Fultz, L.M.*, Davinic, M.*, Acosta-Martinez, V., 2013. Linking soil microbial ecology to ecosystem functioning in integrated crop-livestock systems. Presented at the Special Symposium “Belowground Processes in Grazinglands,” ASA, CSSA, & SSSA International Annual Meeting (Nov 4), Tampa, FL.
Project Outcomes
Our 4th objective was to: translate results from Objectives 1 through 3 into practices incorporated in agriculture in the THP and similar ecosystems. Specifically, we will increase producer and consultant awareness regarding the direct and indirect positive effects for managing agricultural lands to achieve enhanced soil functioning. Our conceptual model is depicted in (Fig 10). Specifically, we increased awareness of the continued direct and positive effects for managing agricultural lands to achieve enhanced soil functioning by the following outreach:
1) Researchers, extension personnel, graduate and undergraduate students attended Dr. a special session at the ASA-CSSA-SSSA meeting, Cincinnati, OH. 21-24 October 2012, entitled, “Integrating Livestock Into Cropping Systems: Ecosystem Responses From Long-Term Studies.” This symposium was organized by Dr. Moore-Kucera and Dr. Fultz (former Ph.D. student under Dr. Moore-Kucera). Dr. Moore-Kucera and her (former) graduate students were co-authors on four presentations highlighted during this session. A total of 11 presentations were given including the keynote address by Dr. Vivien Allen. A list and link to all speakers and abstracts can be found at https://scisoc.confex.com/crops/2012am/webprogram/Session10354.html
The description for this special session was: “Managing agricultural production systems for enhanced ecosystem services such as the sequestration of carbon (C) to offset increasing greenhouse gas (GHG) emissions and enhanced biodiversity and water conservation is a critical focus of agriculture in the US and worldwide. Integrated crop-livestock systems in semi-arid and humid climates have shown promise for conservation of water, nutrients, soil organic C and increased microbial communities. This symposium was a summary of the long-term research conducted in integrated crop-forage-livestock agroecosystems and their impacts on soil-plant-animal interactions and the economic costs and benefits.”
Speakers included: Dr. Vivien Allen, Dr. Veronica Acosta-Martinez, Dr. Ted Zobek, Dr. Lisa Fultz (former Ph.D. student with Dr. Kucera), Dr. Francisco Calderon, Dr. Marko Davinic (former Ph.D. student with Dr. Kucera), Dr. Cody Zilverberg (Dr. Kucera served on Ph.D. committee), Dr. Alan Franzluebbers, Dr. Matt Sanderson, and Dr. Nithya Rajan. This symposium also was attended by Candace Pollock (SSARE public relations coordinator) and highlighted on the SSARE facebook page (October 23, 2012) (Fig 11). https://www.facebook.com/photo.php?fbid=500022700016131&set=a.191931900825214.49340.188127924538945&type=1&theater
2) These interactions during the ASA-CSSA-SSSA meeting eventually resulted in our work being featured on the cover of the CSA News Magazine (58:4-10). The cover article was entitled, “Tiny indicators of change: Microbes in the Texas High Plains may lead to better understanding of how changes in production, environment affect soil health,” written by Madeline Fisher. CSA News Magazine 58:4-10. https://www.soils.org/publications/csa/pdfs/58/8/4 (accessed 8/3/2013).
3) Dr. Moore-Kucera was invited to give a seminar in March 2012 for the College of Life Sciences and Agriculture’s Spring 2012 Sustainable Agriculture and Food Systems Seminar Series at the University of New Hampshire. The title of her seminar was, “Soil microbes and C dynamics under integrated crop-livestock agroecosystems.” The seminar was attended by faculty, college administrators, graduate and undergraduate students and a few local producers.
4) Dr. Moore-Kucera was invited for the 2013 ASA-CSSA-SSSA meeting, held in Tampa, FL to participate in the special symposium, “Belowground Processes in Grazinglands.” The title of this oral presentation was, “Linking soil microbial ecology to ecosystem functioning in integrated crop-livestock systems.” The abstract can be found at https://scisoc.confex.com/crops/2013am/webprogram/Paper77762.html
5) Dr. Moore-Kucera also was invited to speak at a Winter Forages-Livestock Conference held in Blackstone, VA in January 2014. Unfortunately, a family emergency prevented her from attending. She continues to correspond with the organizers to potentially speak at a conference in the future on Soil Microbes and Soil Health: Assessing the Impact of Management on Soil Microbial Communities.
6) To date, 4 peer-reviewed manuscripts have been published and 2 more are in preparation for submission.
7) Two Ph.D. students successfully graduated under this program and both have been hired in career track positions. Dr. Marko Davinic is the R & D Coordinator for Western Ag Innovations Inc and the General Manager for Western Ag Professional Agronomy ND. Dr. Davinic has recently been highlighted in the SSSA publication, Soil Horizons. Dr. Lisa Fultz served 1.5 years as a post-doctoral scientist with Moore-Kucera and will begin her new position as a Soil Scientist at Louisiana State University in Baton Rouge, LA in April 2014.
8) A total of 16 oral or poster presentations have been given at national scientific meetings by PIs and graduate students involved with this project.
As the need for sustainable resources increases, availability of arable land decreases, and levels of atmospheric C increase, these diverse ICL agroecosystems, which have been shown to be beneficial both economically and ecologically, also have the potential to reverse soil degradation in these drought prone climates and may serve as significant SOC accumulators.
Dr. Moore-Kucera continues to collaborate with scientists and producers. She is currently collaborating with other TTU researchers, who are are actively pursuing further funding to continue to identify agricultural management strategies that enhance ecosystem services while conserving resources and supporting economic growth.
- Fig. 10. Conceptual framework that depicts the major ecological benefits and mechanisms that impart these benefits as a result of integrated crop-livestock (ICL) agroecosystems in comparison to continuous cotton cropping systems (CTNc).
- Fig. 11. Some of the invited speakers of the special ASA-CSSA-SSSA session organized by Moore-Kucera and Fultz entitled, “Integrating Livestock Into Cropping Systems: Ecosystem Responses From Long-Term Studies.”
{kind=link}
Economic Analysis
While economic analysis was not the primary focus of this research, Allen et al (2008), Allen et al. (2012), Zilverberg et al. (2012) discussed the economic ramifications associated with systems examined for this research.
Farmer Adoption
Dr. Moore-Kucera continues to collaborate with scientists and producers working in ICL agroecosystems. She is currently collaborating with other TTU researchers, who are actively pursuing further funding to continue to identify agricultural management strategies that enhance ecosystem services while conserving resources and supporting economic growth. We hope our results add another layer of ecological benefits imparted by ICL agroecosystems in the Southern High Plains region and other similar areas. We continue to work closely with the Texas Alliance for Water Conservation producers (over 26 producer-operated farms in the SHP region aimed at water conservation; many of these systems are ICLs). We hope that our results that support our conceptual framework (Fig 10) for enhanced ecosystem sustainability will provide farmers and land managers critical information to drive management decisions.
Basically our conceptual framework resulting from ICL management is that systems that support low-disturbance and relatively high vegetative residues to the soil will help build soil organic matter, increase microbial biomass, diversity, and activity which together help to cycle nutrients and stabilize and form macroaggregates that are important for water storage and protection from erosion. Additionally, the stabilization of macroaggregates can physically protect soil C in smaller aggregates inside of these larger macroaggregates, which can help to sequester C.
What our results suggested was that there was a gradient in management options that supported enhanced properties associated with sustainability. This is very important because producers have options. For example, as water resources in our region become more and more limited, producers can choose how they transition to new systems. Dryland or deficit irrigated ICL systems can provide some flexibility as producers learn about forages and cattle management if they were formerly producing 100% annual crops; alternatively mixing some land under dryland perennial forages and some under irrigated crops or forages also helps during times of drought to make a crop. The goal is to approach sustainability from ecological and socioeconomic levels to provide the flexibility and minimize risks.
Areas needing additional study
Areas that remain unclear include the response of the ICL ecosystems to climate change including the future predictions of intense and more frequent drought. Our region is currently in a long-standing drought. How the microbial communities in conjunction with drought-tolerant vegetation can facilitate ecosystem resiliency during these challenging climate patterns will be necessary to sustain agriculture in our region.
Another area that needs additional study is a more in-depth greenhouse gas flux study that specifically targets N2O. Our one year of data (i.e., before the worst drought on record began in 2011), suggested that intensively N fertilized bermudagrass (relative to the other crops) may only partially offset and not fully compensate for the increased organic C sequestration potential. However, N2O fluxes are highly variable. We hypothesize that systems that add N fertilizers via subsurface drip systems may lower the rate of release of N2O due to diffusion limitations in the soil. Better timing and delivery of N fertilizers, regardless of the crop will help minimize the release of this potent greenhouse gas.
Literature cited
- Acosta-Martínez, V., Bell, C.W., Morris, B.E.L., Zak, J., Allen, V.G., 2010. Long-term soil microbial community and enzyme activity responses to an integrated cropping-livestock system in a semi-arid region. Agriculture, Ecosystems and Environment 137, 231-240.
- Acosta-Martínez, V., Zobeck, T.M., Gill, T.E., Kennedy, A.C., 2003. Enzyme activities and microbial community structure in semiarid agricultural soils. Biology and Fertility of Soils 38, 216-227.
- Ball, B.C., Scott, A., Parker, J.P., 1999. Field N2O, CO2, and CH4 fluxes in relation to tillage, compaction and soil quality in Scotland. Soil and Tillage Research 53, 29-36.
- Cambardella, C.A., Elliott, E.T., 1993. Carbon and nitrogen distribution in aggregates from cultivated and native grassland soils. Soil Sci Soc Am J 57, 1071-1076.
- Cambardella, C.A., Elliott, E.T., 1994. Carbon and nitrogen dynamics of soil organic matter fractions from cultivated grassland soils. Soil Sci Soc Am J 58, 123-130.
- Cerri, C.E.P., Coleman, K., Jenkinson, D.S., Bernoux, M., Victoria, R., Cerri, C.C., 2003. Modeling soil carbon from forest and pasture ecosystems of Amazon, Brazil. Soil Sci Soc Am J 67, 1879-1887.
- Davinic, M., Fultz, L.M., Acosta-Martinez, V., Calderón, F.J., Cox, S.B., Dowd, S.E., Allen, V.G., Zak, J.C., Moore-Kucera, J., 2012. Pyrosequencing and mid-infrared spectroscopy reveal distinct aggregate stratification of soil bacterial communities and organic matter composition. Soil Biology and Biochemistry 46, 63-72.
- Davinic, M., Moore-Kucera, J., Acosta-Martínez, V., Zak, J., Allen, V., 2013. Soil fungal distribution and functionality as affected by grazing and vegetation components of integrated crop-livestock agroecosystems. Applied Soil Ecology 66, 61-70.
- Denef, K., Zotarelli, L., Boddey, R.M., Six, J., 2007. Microaggregate-associated carbon as a diagnostic fraction for management-induced changes in soil organic carbon in two Oxisols. Soil Biology and Biochemistry 39, 1165-1172.
- Dobranic, J.K., Zak, J.C., 1999. A microtiter plate procedure for evaluating fungal functional diversity. Mycologia 91, 756-765.
- Frostegård, A., Bååth, E., 1996. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biology and Fertility of Soils 22, 59-65.
- Frostegård, Å., Tunlid, A., Bååth, E., 2011. Use and misuse of PLFA measurements in soils. Soil Biology and Biochemistry 43, 1621-1625.
- Fultz, L.M., Moore-Kucera, J., Zobeck, T.M., Acosta-Martinez, V., Allen, V.G., 2013a. Aggregate Carbon Pools after 13 Years of Integrated Crop-Livestock Management in Semiarid Soils. Soil Sci Soc Am J 77, 1659-1666.
- Fultz, L.M., Moore-Kucera, J., Zobeck, T.M., Acosta-Martínez, V., Wester, D.B., Allen, V.G., 2013b. Organic carbon dynamics and soil stability in five semiarid agroecosystems. Agriculture, Ecosystems & Environment 181, 231-240.
- Graham, J.H., Hodge, N.C., Morton, J.B., 1995. Fatty acid methyl ester profiles for characterization of glomalean fungi and their endomycorrhizae. Applied and environmental microbiology 61, 58-64.
- Hutchinson, G.L., Mosier, A.R., 1981. Improved soil cover method for field measurement of nitrous oxide fluxes. Soil Science Society of America Journal 45, 311-316.
- Kong, A.Y.Y., Six, J., Bryant, D.C., Denison, R.F., Van Kessel, C., 2005. The relationship between carbon input, aggregation, and soil organic carbon stabilization in sustainable cropping systems. Soil Sci Soc Am J 69, 1078-1085.
- Lal, R., 2004. Soil carbon sequestration impacts on global climate change and food security. Science 304, 1623-1627.
- Lal, R., 2009. Challenges and opportunities in soil organic matter research. Eur J Soil Sci 60, 158-169.
- Lal, R., Follet, R.F., 2009. Soils and climate change, in: Lal, R., Follet, R.F. (Eds.), Soil Carbon Sequestration and the Greenhouse Effect. SSSA Special Publication, Madison, WI.
- Madan, R., Pankhurst, C., Hawke, B., Smith, S., 2002. Use of fatty acids for identification of AM fungi and estimation of the biomass of AM spores in soil. Soil Biology and Biochemistry 34, 125-128.
- Magurran, A.E., 1988. Ecological Diversity and its Measurement. Princeton University Press, New Jersey.
- Olsson, P.A., 1999. Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil. FEMS microbiology ecology 29, 303-310.
- Schutter, M.E., Dick, R.P., 2000. Comparison of fatty acid methyl ester (FAME) methods for characterizing microbial communities. Soil Sci Soc Am J 64, 1659-1668.
- Six, J., Elliott, E.T., Paustian, K., 1999. Aggregate and soil organic matter dynamics under conventional and no-tillage systems. Soil Sci Soc Am J 63, 1350-1358.
- Six, J., Elliott, E.T., Paustian, K., 2000. Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture. Soil Biology and Biochemistry 32, 2099-2103.
- Six, J., Elliott, E.T., Paustian, K., Doran, J.W., 1998. Aggregation and soil organic matter accumulation in cultivated and native grassland soils. Soil Sci Soc Am J 62, 1367-1377.
- Sobek, E.A., Zak, J.C., 2003. The Soil FungiLog procedure: Method and analytical approaches toward understanding fungal functional diversity. Mycologia 95, 590-602.
- Tabatabai, M.A., 1994. Soil enzymes. Methods of Soil Analysis. Part 2. Microbiological and Biochemical Properties, 775-833.
- Zak, J.C., Willig, M.R., Moorhead, D.L., Wildman, H.G., 1994. Functional diversity of microbial communities: A quantitative approach. Soil Biology and Biochemistry 26, 1101-1108.
- Zelles, L., 1997. Phospholipid fatty acid profiles in selected members of soil microbial communities. Chemosphere 35, 275-294.
- Zelles, L., Adrian, P., Bai, Q.Y., Stepper, K., Adrian, M.V., Fischer, K., Maier, A., Ziegler, A., 1991. Microbial activity measured in soils stored under different temperature and humidity conditions. Soil Biology and Biochemistry 23, 955-962.
Information Products
- Pyrosequencing and mid-infrared spectroscopy techniques reveal distinct aggregate stratification of soil bacterial communities and organic matter composition.
- Soil fungal distribution and functionality as affected by grazing and vegetation components of integrated crop-livestock agroecosystems
- Aggregate carbon pools after 13-years of integrated crop-livestock management in semi-arid soils
- Organic carbon dynamics and soil stability in five semiarid agroecosystems